L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
Moscamed.org.br
Fruit Flies of Economic Importance: From Basic to Applied Knowledge
Proceedings of the 7th International Symposium on Fruit Flies of Economic Importance
new genetic tools for Improving SIt in Ceratitis capitata: Embryonic lethality and Sperm Marking
Marc F. Schetelig1, Francesca Scolari2, Alfred M. Handler3,
1 Department of Developmental Biology, Göttingen Center for Molecular Biosciences, Johann-Friedrich Blumenbach Institute of Zoology and Anthropology, Georg-August-University Göttingen, Justus-von-Liebig-Weg 11, 37077 Göttingen, Germany. 2 Dipartimento di Biologia Animale, Università di Pavia, Piazza Botta 9, 27100 Pavia, Italy. 3 USDA/ARS, Center for Medical, Agricultural and Veterinary Entomology, Gainesville, Florida, USA.ABStrAct: Environment friendly sterile insect technique (SIT) is being applied effectively as a component of area-wide integrated pest management (AW-IPM) for Ceratitis capitata since 1970s. Nevertheless improved biological strategies are needed to increase the efficacy of AW-IPM. Transgenic approaches should increase and widen the applicability of such programmes to different pest species. In this respect two major strategies are followed: First an approach to cause sterility was designed without interfering with spermatogenesis to maintain males and their sperm as competitive as possible. We followed a strategy, which is based on the expression of a lethal factor under the control of a promoter that is active at early blastoderm stages. The system employs the ectopic expression of a hyperactive proapoptotic gene that causes embryo-specific lethality when driven by the tetracycline-controlled transactivator tTA under the regulation of a cellularization gene enhancer/promoter. The system has been tested successfully in Drosophila melanogaster (Horn & Wimmer 2003). We tried the direct transfer of the Drosophila system to Ceratitis capitata by injecting the respective constructs that carry Drosophila-derived promoters. Unfortunately, the cellularization specific promoters from Drosophila seem not functional in Ceratitis. Therefore, the corresponding enhancers/promoters from Ceratitis were isolated and subsequently the tTA was brought independently under the control of each enhancer/promoter region. These constructs were injected in Ceratitis for further evaluation. Second, we have engineered a medfly strain carrying a sperm marking system. This strain carries two fluorescent markers. One (turboGFP) marker is under the control of the spermatogenesis specific b2-tubulin promoter from Ceratitis and is therefore sperm specifically expressed. The second (DsRed) is under the control of the polyubiquitin promoter of Drosophila. Released males from this strain could be distinguished from wildtype males in the monitoring process. In addition, monitoring of the mating success of released sterile and fluorescently sperm-marked males by trapping females and examine their spermathecae would be possible. This effective and easily recognizable sperm marking will make novel studies possible on medfly reproductive biology and using sperm marked strains could optimize releasing strategies in SIT-based AW-IPM.
Key Words: cellularization, conditional embryonic lethality, insect transgenesis, sperm marker, beta2-tubulin. IntroductIon
vegetables and nuts) and on the ability to adapt its multivoltine cycle to different tem-
The Mediterranean fruit fly, Ceratitis capi-
perate climates to overwinter as larvae in
tata (Wiedemann) (Diptera: Tephritidae), is
Biological approaches to insect pest man-
agement offer alternatives to insecticidal
species belonging to the Tephritidae family
control. The Sterile Insect Technique (SIT)
(Khoo et al. 1991). A native of Sub-Saharan
is regarded as an ecologically safe method
Africa, medfly has spread into the Mediter-
for area-wide control. SIT reduces the pest
ranean basin to parts of Central and South
population by mass release of sterilized pest
America, Hawaii and Australia in less than
organisms (Knipling 1955). The SIT process
200 years. Moreover, in the latter half of the
involves mass rearing and release of indi-
last century, it was sporadically detected
vidual flies rendered sterile through gamma
in different areas of the United States like
or X-ray irradiation exposure. SIT has been
California, Florida and Texas. The success of
proven in many areas to be effective against
C. capitata infestation is partially dependant
on its highly polyphagous nature (C. capi-tata attacks more than 250 different fruits,
tion of C. capitata was achieved over a region of 15.000 km2 in 1982 (Hendrichs et al. 1983)
mala (Villasenor et al. 2000). For many pest
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
species including medfly, SIT proved to be
based on the use of the C. capitata sper-
most effective by releasing only sterile
matogenesis-specific b2t promoter driving
a fluorescent marker (Scolari et al. 2008).
of radiation for sterilizing insects does have
After thorough strain evaluation and a test
some adverse effects on their competitive-
phase for fitness, accuracy and stability of
ness which in turn reduces the efficiency of
the sexing procedure as well as the stabil-
the technique (Bushland 1971; Cayol et al.
ity of these strains, they could be used for
different purposes. A possible application
onic sterility system was successfully estab-
lished in Drosophila melanogaster (Meigen)
strain in combination with the ability for an
easy monitoring in an operational SIT pro-
may provide an alternative to the use of ra-
gram. The system will also help in providing
diation in area-wide integrated pest man-
more detailed information on reproductive
biology of C. capitata. The aim of the stud-
ies reported here was to establish and eval-
grams, mass reared pupae are sterilized and
marking systems in C. capitata. Functional
large-scale SIT activities, like those estab-
enable sterilized flies to be distinguished
lished for C. capitata, are ideal for compar-
from wild flies when recaptured in traps
in the release area. This monitoring system
implies some disadvantages: the fluores-cent dyes are expensive, dangerous for hu-man health and error prone, because they
MAtErIAl And MEthodS
can be transferred from sterilized marked flies to wild type ones (Hagler and Jack-
In-situhybridization. The RNA probes were
made with a DIG-RNA-labeling Kit (Roche,
monitoring in various species. A dominant
formed as described in Davis et al. (2001).
mutation has been isolated for C. capi-Plasmid. pB[s1-tTA; PUb-DsRed1] was con-
tata, which affects the third stripe on the
structed as previously described in Schetelig
abdomen (Niyazi et al. 2005). Field-cage
studies have shown comparable sexual compatibility and mating competitiveness, including data on genetic sexing strains
(GSS). Another system describes a trans-genic sexing strain in Anopheles stephensi
1) Evaluation of a transgenic approach to
(Catteruccia et al. 2005). In this system the
sterilize flies with an embryonic lethal trans-
spermatogenesis specific b2-tubulin (b2t)
gene combination in C.capitata.
promoter from Anophelesgambiae drives
An approach to cause sterility was designed
enhanced green fluorescent protein (EGFP).
without interfering with spermatogenesis to
maintain males and their sperm as competi-
scale conditions as a good sexing strain in
tive as possible. We followed a strategy based
on the expression of a lethal factor under the
control of a promoter that is active at early
marking system for C. capitata, which is
blastoderm stages. When the male is homozy-
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
gous for the combination of the necessary
for endogenous promoters of C. capitata. To
gene constructs, each fertilization event will
obtain C. capitata genes and their promoters
lead to embryonic lethality (Horn and Wim-
which lead to specific expression at the blas-
mer 2003). The advantage of this system lies
toderm stage, we first carried out PCR-based
cDNA subtractions of different embryonic
such males, since their reproductive organs
stages and identified several cellularization-
will not be affected and matings actually lead
specific genes (Schetelig et al. 2007). After
to sperm transfer. However, it is very impor-
that we isolated the corresponding enhanc-
tant that the promoter is active only in early
stages of development. Then the lethal phase
dently under the control of each enhancer/
permissive conditions in the rearing facilities,
promoter region. We injected the construct
whereas after release non-permissive con-
carrying the cellularization specific promoter
ditions will not affect the males themselves
of sub1_68 from C. capitata into the germline
but only their progeny (Wimmer 2005). The
of C. capitata and got transgenics with cel-
system employs the ectopic expression of a
lularization specific expression of tTA (Fig-
hyperactive proapoptotic gene that causes
ure 2). However, the expression as detected
embryo-specific lethality when driven by
by whole mount in-situ hybridizations was
the tetracycline-controlled transactivator
tTA under the regulation of a cellularization
marking system for Ceratitis capitata.
been tested successfully in Drosophila mela-
To develop a sperm marking system for the
nogaster (Horn and Wimmer 2003). We want
fruit fly C. capitata, we isolated the spermato-
to transfer this binary expression system to
genesis specific gene b2t from C. capitata by
C. capitata to evaluate it in comparison with
degenerate primer PCR. By rapid amplifica-
the effective conventional SIT using radia-
tion of cDNA ends (RACE), we amplified the
complete b2t and isolated the upstream
We first tried the direct transfer of the Droso-
region by iPCR. The observed upstream re-
phila-derived system to C. capitata. Therefore
gion of b2t was fused to the fluorescent
we injected the driver construct pB[s1-tTA;
marker turboGFP (tGFP). b2t-tGFP was then
PUb-DsRed1], which contains the tTA gene
inserted in a piggyBac (pB) transposon vec-
under the control of the Drosophila melano-
tor carrying a red (DsRed) fluorescent marker
gaster serendipity a (srya) promoter region
driven by a polyubiquitin promoter (PUb):
(Schetelig et al. 2007), together with the
b2t-tGFP in pB[PUb-DsRed1]. The advantage
phsp-pBac (Handler and Harrell 1999) into
of this transposon vector is that it carries a
the germline of C. capitata. We got four inde-
characterized fluorescent marker driven by
pendent transgenic lines. These lines where
the ubiquitiously expressed polyubiquitin
tested for tTA expression by whole mount in-
promoter from Drosophila melanogastersitu hybridizations with a RNA probe to tTA.
(Handler and Harrell 2001). The construct
None of the four transgenic lines expressed
pB[b2-tGFP; PUb-DsRed1] was injected with
the tTA. Representative three of the four lines
a transposase source into the posterior end
of C. capitata embryos to cause germline
Thus, the cellularization specific srya pro-
transformation. Flies from this transforma-
moter from Drosophila seems not to be func-
tion have a red body and males have green
tional in C. capitata. In order to get functional
fluorescent testes (Figure 3). For sperm use
promoters for use in our system, we searched
and remating analyses different fluorescent
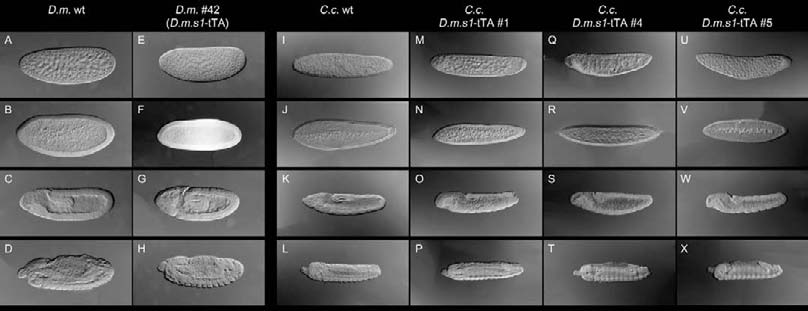
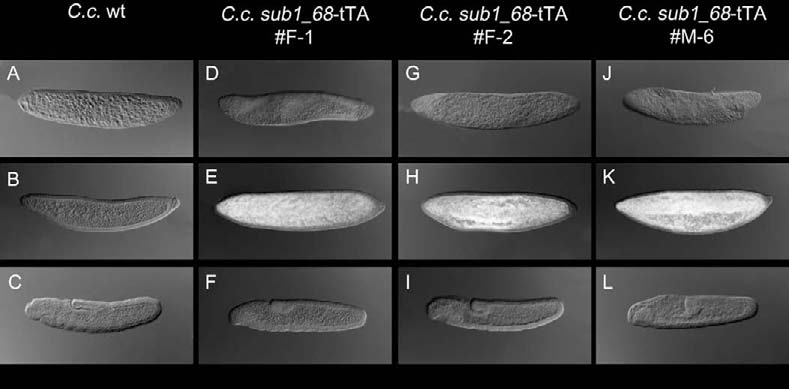
Schetelig et al.: Embryonic lethality and sperm marking in CeratitisFig. 1. Cellularization specific promoter of Drosophila is apparently nonfunctional in C. capitata. Shown are whole mount in-situ hybridizations using a tTA RNA probe. The embryogenesis of each strain (columns) is pictured with four embryonic stages: first row = blastoderm; second row = cellularization; third row = germ band elongation; fourth row = germ band retraction. D.m. wildtype (A-D): no expression of tTA at all stages. D.m. #42 (Horn and Wimmer 2003) (E-H): tTA is cellularization specifically expressed in this strain, which carries the D.m.s1-tTA. C.c. wildtype (I-L): no expression of tTA at all stages. C.c.D.m.s1-tTA #1 (M-P), C.c.D.m.s1-tTA #4 (Q-T) and C.c.D.m.s1- tTA #5 (U-X): no expression of tTA at all stages. Fig. 2. Endogenous promoter of a cellularization specific gene of C. capitata causes weak tTA expression. Shown are whole mount in-situ hybridizations using a tTA RNA probe. The embryogenesis of each strain (columns) is pictured with three embryonic stages: first row = blastoderm; second = cellularization; third = germ band elongation. C.c. wildtype control (A-C): no expression of tTA at all stages. C.c.sub1_68-tTA #F-1 (D-F), C.c.sub1_68-tTA #F-2 (G-I) and C.c.sub1_68-tTA #M-6 (J-L): cellularization specific expres- sion of tTA driven by the C.c.sub1_68 promoter.
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
marked sperms would be helpful. Therefore
and their promoters. We could show that the
we are developing also a system carrying a
Ceratitis cellularization specific promoter from
sub1_68 fused to tTA expressed tTA specifically at cellularization stages (Figure 2). So this pro-moter might be sufficient for using it for our embryonic lethality system. But the staining time for the color reaction of 24h for the de-picted whole mount in-situ hybridizations was not in an estimated average time window for in-situ hybridizations on C. capitata with RNA probes (average of staining time to the color reaction is completed in C. capitata embryos is between 0.5 and 3h). So we suppose that the specific, albeit low expression of this construct will probably not be sufficient for generating a successful and safe lethality system. To bet-ter promote the tTA expression we wish to isolate varying long versions of the sub1_68 upstream region which might include more
Fig. 3. Transgenic marked C. capitata male carrying two
regulating elements. We will also isolate and
different types of molecular markers. The male has a red
test upstream regions from other C. capitata
fluorescent body marker (PUb-DsRed) and a testes-
specific green fluorescent marker (b2t-tGFP).
2) Development of a transgenic sperm mark-
ing system for Ceratitis capitata.
dIScuSSIon
After thorough strain evaluation and test
phase for fitness, accuracy and stability, the
1) Evaluation of a transgenic approach to
developed sperm marked strains are a first
sterilize flies with an embryonic lethal trans-
step for improving the following objectives
gene combination in Ceratitiscapitata.
A direct transfer of the driver construct carry-
• Reproductive biology in Ceratitis capitata.
ing the cellularization specific D. melanogaster
Thus far little is known about the mecha-
srya promoter to C. capitata showed that this
nisms of remating behavior and the conse-
promoter is not functional in C. capitata. Thus,
we can assume that the complex interaction
ing system it should be possible to widen
between enhancers and promoters of stage-
our the knowledge of the reproductive bi-
specifically expressed genes (Blackwood and
ology of the medfly. The obtained results
Kadonaga 1998) is different between D. mela-
will also be important to further improve
nogaster and C. capitata. A Drosophila melano-gaster promoter might not act as an adequate
• Monitoring of an area wide SIT program.
alternative to an endogenous C. capitata pro-
Because b2t is expressed in the testes, re-
moter to enable strong expression rates.
leased males from these strains could be
For a highly specific embryonic lethality sys-
distinguished from wildtype males in the
expression specific to early embryonic stages.
Therefore we isolated several cellularization
in the field and dissection of their sperm
genes from C. capitata (Schetelig et al. 2007)
storage organs, this would allow for a bet-
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
ter evaluation of the efficiency of SIT pro-
(Diptera: Tephritidae) laboratory-reared strains under
field cage conditions. Journal of Economic Entomol-
• In principle these strains can also be used
Davis, G.K., Jaramillo, C.A., and N.H. Patel. 2001. Pax group
III genes and the evolution of insect pair-rule pat-
tomatically for males and females during
terning. Development 128: 3445-3458.
Hagler, J.R., and C.G. Jackson. 2001. Methods of marking
insects: current techniques and future prospects. An-
since Ceratitis capitata has already an ef-
nual Review of Entomology 46: 511-543.
Handler, A.M., and R.A. Harrell II. 1999. Germline trans-
fective sexing strain, this might be more
formation of Drosophila melanogaster with the pig-gyBac transposon vector. Insect Molecular Biology 8:
Handler, A.M., and R.A. Harrell II. 2001. Polyubiquitin-reg-
ulated DsRed marker for transgenic insects. BioTech-niques 31: 820-828.
AcknoWlEdgEMEntS
Horn, C., and E.A. Wimmer. 2003. A transgene-based,
embryo-specific lethality system for insect pest man-
agement. Nature Biotechnology 21: 64-70.
Khoo, K.C., Ooi, P.A.C., and C.T. Ho. 1991. Crop Pests and
ert Bosch Foundation (EAW) within the pro-
their management in Malaysia. Malaysia: Tropical
gram 'International Research into the De-
velopment of Sustainable Agriculture and
Knipling, E.F. 1955. Possibilities of insect control or eradi-
Forestry', by the Fonds der chemischen In-
cation through the use of sexually sterile males.Jour-
dustrie (EAW), by the Italian Ministry of
nal of Economic Entomology 48: 459-462.
Hendrichs, J., Ortiz, G., Liedo, P., and A. Schwarz. 1983. Six
Education, University & Research within
years of successful medfly program in Mexico and
Guatemala, pp 353-365. In R. Cavalloro (ed.), Proceed-
di Ricerca Scientifica di Rilevante Interesse
ings, Symposium: Fruit Flies of Economic Importance.
Nazionale; GG), by the University of Pavia
CEO/IOBC International Symposium, 16-19 Novem-
within the program 'FAR 2005' (Fondi di
ber 1982, Athens, Greece. A.A. Balkema, Rotterdam,
Ateneo-Ricerca; GG) and the USDA National
Niyazi, N., Caceres, C., Delprat, A., Wornoayporn, V., Rami-
Research Initiative Competitive Grants Pro-
rez Santos, E., Franz, G., and A.S. Robinson. 2005. Ge-
netics and mating competitiveness of Ceratitis capi-tata (Diptera: Tephritidae) strains carrying the marker Sergeant, Sr 2. Annals of the Entomological Society of America 98: 119-125
rEFErEncES
Robinson, A.S. 2002. Genetic sexing strains in medfly, Cer-atitis capitata, sterile insect technique programmes.
Blackwood, E.M., and J.T. Kadonaga. 1998. Going the dis-
tance: a current view of enhancer action. Science 281:
Schetelig, M.F., Horn, C., Handler, A.M., and E.A. Wimmer.
2007. Development of an embryonic lethality system
Bushland, R.C. 1971. Sterility principle for insect control.
for SIT in Ceratitis capitata. InArea-wide Control of
In: Historical development and recent innovations
Insect Pests: From Research to Field Implementation.
in sterility principle for insect control or eradication.
M.J.B. Vreysen, A.S. Robinson and J. Hendrichs (eds.).
Proceedings of a consultants meeting, IAEA, Vienna.
Springer, Dordrecht, The Netherlands, pp. 85-93.
Scolari, F., Schetelig, M.F., Bertin, S., Malacrida, A.R., Gas-
Catteruccia, F., Benton, P.J., and A. Crisanti. 2005. An
peri, G. & Wimmer, E.A. 2008. Fluorescent Sperm Mark-
Anopheles transgenic sexing strain for vector control.
ing to Improve the Fight against the Pest Insect Cera-titis capitata (Wiedemann; Diptera: Tephritidae). New
Cayol, J.P., Vilardi, J., Rial, E., and M.T. Vera. 1999. New in-
dices and method to measure the sexual compat-
Villasenor, A., Carrillo, J., Zavala, J., Stewart, J., Lira, C., and
ibility and mating performance of Ceratitis capitata
J. Reyes. 2000. Current progress in the medfly pro-
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
gram Mexico-Guatemala, pp. 361-368. In K.H. Tan
Importance, 28 May-5 June 1998, Penang, Malaysia.
(ed.), Proceedings: Area-Wide Control of Fruit Flies
Penerbit Universiti Sains Malaysia, Pulau Pinang, Ma-
and Other Insect Pests. International Conference
on Area-Wide Control of Insect Pests, and the 5th In-
Wimmer, E.A. 2005. Eco-friendly insect management. Na-
ternational Symposium on Fruit Flies of Economic
06 Normas y congreso.qxd 21/7/08 10:45 Página 27El presente documento es una adaptación de la «Guía para los Autores» del Journal of the American Dietetic Association 1 y los estándares de publicaciónde Doyma-Elsevier elaborada por el Comité Editorial de ACTIVIDAD DIETÉTICA. La revista ACTIVIDAD DIETÉTICA es el órgano oficial de expresión de la Asociación Española de Dietistas-Nut
 Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
Fig. 1. Cellularization specific promoter of Drosophila is apparently nonfunctional in C. capitata. Shown are whole
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
Fig. 1. Cellularization specific promoter of Drosophila is apparently nonfunctional in C. capitata. Shown are whole 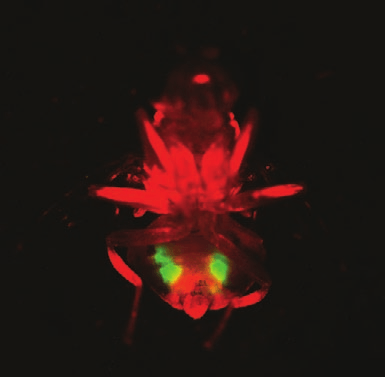 Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
marked sperms would be helpful. Therefore
and their promoters. We could show that the
we are developing also a system carrying a
Ceratitis cellularization specific promoter from
sub1_68 fused to tTA expressed tTA specifically at cellularization stages (Figure 2). So this pro-moter might be sufficient for using it for our embryonic lethality system. But the staining time for the color reaction of 24h for the de-picted whole mount in-situ hybridizations was not in an estimated average time window for in-situ hybridizations on C. capitata with RNA probes (average of staining time to the color reaction is completed in C. capitata embryos is between 0.5 and 3h). So we suppose that the specific, albeit low expression of this construct will probably not be sufficient for generating a successful and safe lethality system. To bet-ter promote the tTA expression we wish to isolate varying long versions of the sub1_68 upstream region which might include more
Fig. 3. Transgenic marked C. capitata male carrying two
Schetelig et al.: Embryonic lethality and sperm marking in Ceratitis
marked sperms would be helpful. Therefore
and their promoters. We could show that the
we are developing also a system carrying a
Ceratitis cellularization specific promoter from
sub1_68 fused to tTA expressed tTA specifically at cellularization stages (Figure 2). So this pro-moter might be sufficient for using it for our embryonic lethality system. But the staining time for the color reaction of 24h for the de-picted whole mount in-situ hybridizations was not in an estimated average time window for in-situ hybridizations on C. capitata with RNA probes (average of staining time to the color reaction is completed in C. capitata embryos is between 0.5 and 3h). So we suppose that the specific, albeit low expression of this construct will probably not be sufficient for generating a successful and safe lethality system. To bet-ter promote the tTA expression we wish to isolate varying long versions of the sub1_68 upstream region which might include more
Fig. 3. Transgenic marked C. capitata male carrying two