L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
Leishmania donovani possess a nadph-dependent alkylglycerol cleavage enzyme
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS 227, 885 - 889 (1996) Leishmania donovani Possess a NADPH-Dependent
Deqin Ma,* Stephen M. Beverley,† and Salvatore J. Turco*,1
*Department of Biochemistry, University of Kentucky Medical Center, Lexington, Kentucky 40536; and
†Department of Biological Chemistry and Molecular Pharmacology,Harvard Medical School, Boston, Massachusetts 02115Leishmania parasites possess an abundance of ether-linked hydrocarbons as components of phospholipids
and glycosylphosphatidylinositol anchors of glycoproteins and polysaccharides, including important surfacemolecules such as lipophosphoglycan (LPG) and glycosylinositolphospholipids (GIPLs). Cleavage of theether bond is an important feature in the turnover pathway of alkylglycerols. In mammals, ether lipidcleavage activity requires a pteridine cofactor (H4-biopterin), suggesting the potential for linkage betweenthe unusual Leishmania pteridine metabolic pathways and lipid metabolism. In this study, we partiallypurified and characterized an activity in L. donovani capable of cleaving the ether lipid 1-O-alkyl[3H]glycol. Unlike the mammalian enzyme but like that of Tetrahymena, the Leishmania enzyme required NADPHrather than H4-biopterin. The use of divergent cofactors by the parasite and mammalian enzymes mayprovide a basis for the design of anti-parasitic drugs targeting ether-linked lipid metabolism.
Key determinants of Leishmania infectivity and survival in hostile environments of the host
are its surface glycoconjugates lipophosphoglycan (LPG) and glycosylinositolphospholipids(GIPLs). LPG is a polymorphic glycosylphosphatidylinositol (GPI)-anchored polysaccharide. The lipid portion of LPG characteristically contains a 1-O-alkyl-2-lyso-phosphatidylinositol inwhich the aliphatic chain consists of either a C-24 or C-26 saturated, unbranched hydrocarbon(1,2). While all of the GIPLs have the identical 1-O-alkylglycerol moiety, some possess anesterified fatty acid on C-2 of the glycerol backbone.
In mammalian cells, the turnover of 1-O-alkylglycerol requires a glyceryl ether monooxygen-
ase (1-O-alkyl-sn-glycerol, tetrahydropteridine:oxygen oxidoreductase, EC 1.14.16.5) that cata-lyzes the oxidative cleavage of the lipid to fatty aldehyde and glycerol (3). The monooxygenasecan likewise degrade alkylglycol. This reaction requires molecular oxygen and tetrahydrobio-pterin to hydroxylate the aliphatic carbon adjacent to the ether bond. The product of the initialreaction is a hemiacetal which spontaneously hydrolyzes to yield the fatty aldehyde andfree glycerol. Relatively low concentration of alkylglycerol can be efficiently catabolized byLeishmania whereas high concentrations (§ 25 mM) of alkylglycerol are toxic (4). Due tothe abundance of the alkylglycerol-anchored LPG and GIPLs on the surface of Leishmaniapromastigotes and the toxicity of alkylglycerol to the parasites, we investigated the nature ofthe parasitic enzyme that degrades alkylglycerol. In this paper, we report the discovery of anenzyme in Leishmania capable of catalyzing the cleavage of alkylglycol. The enzyme was
1 To whom correspondence should be addressed. Fax: 606-323-1037. This investigation received financial
support from NIH Grants AI20941 and AI31078. S.M.B. and S.J.T. are Burroughs Wellcome Scholars in MolecularParasitology.
Abbreviations: lipophosphoglycan (LPG); glycosylinositolphospholipid (GIPL); glycosylphosphatidylinositol (GPI);
tetrahydrobiopterin (BPTH4); pteridine reductase (PTR1).
0006-291X/96 $18.00Copyright ᭧ 1996 by Academic Press, Inc. All rights of reproduction in any form reserved.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
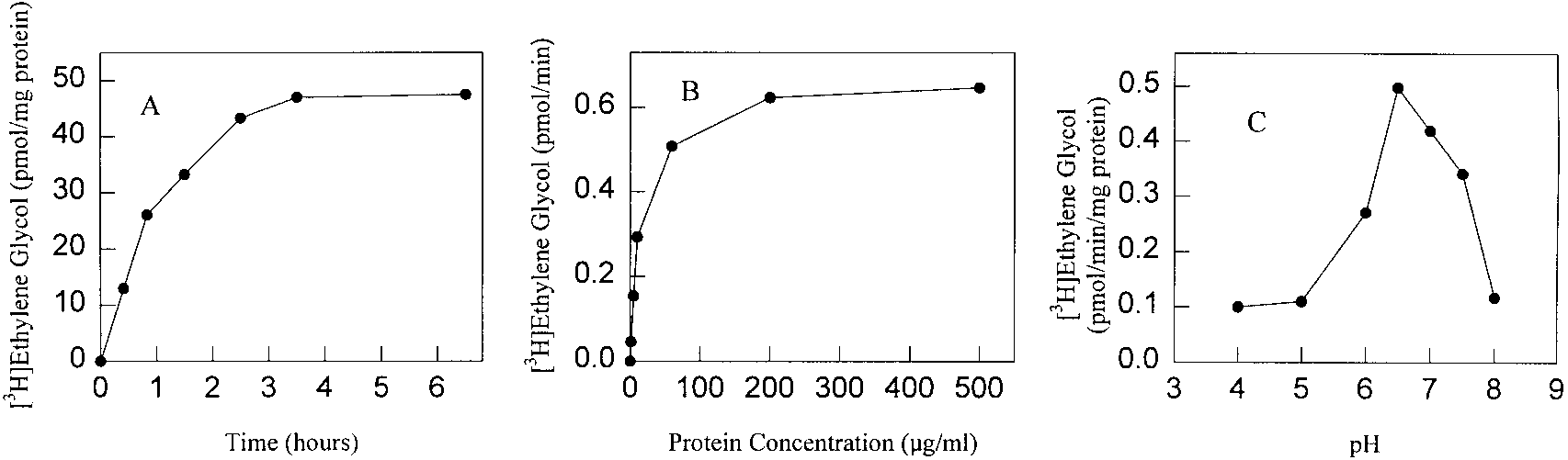
FIG. 1. Analysis of alkylglycerol cleavage activity in a cell-free system of L. donovani. Membranes (2 mg/ml)
were incubated with 0.01 mCi of 1-O-octadecyl[3H]glycol at 35ЊC for 3.5 h unless otherwise noted. At the appropriatetime, aliquots of the reaction mixture were removed and extracted as described in Materials and Methods. A, timecourse. B, protein concentration. C, pH curve.
partially purified and characterized and found to require NADPH as a coenzyme unlike itsmammalian counterpart. Materials. All materials were obtained as follows: Dulbecco's modified Eagle's medium (DME 430-2100) and fetal
calf serum from Gibco; Bacto-Brain Heart Infusion (0037-05-2) from Difco; 2-amino-6,7-dimethyl-4-hydroxy-5,6,7,8-tetrahydropteridine hydroxychloride from Aldrich Chem. Co., 1-O-octyldecylglycerol, QAE-cellulose, NADPH, cata-lase, and glutathione from Sigma, (6R) 5,6,7,8,-tetrahydrobiopterin from RBI. Parasites. Promastigotes of L. donovani were maintained as described elsewhere (5). For isolation and partial
purification of enzyme, one-liter cultures of parasites were grown in Brain-Heart Infusion (6). Preparation of 1-O-octadecyl[3H]glycol. 1-O-octadecyl[3H]glycol was prepared by a modification of the procedure
of Kaufman et al. (7). Batyl alcohol (25 mg) was dissolved at 5 mg/ml in 90% acetic acid containing 50 mM sodiumperiodate. After a 1 hour incubation at room temperature in darkness, 5 ml of water was added and the reactionmixture was extracted twice with 1 vol. hexane/diethyl ether (1:1, v/v). The upper phase was dried under nitrogen,redissolved in absolute ethanol, and transferred to a tube containing 5 mCi of NaB[3H]4. After a 12 hour incubationat room temperature, 25 mg NaBH4 was added to the solution and then incubated for another 2 hours. One volumeof 0.5 M HCl was then added and the mixture was extracted twice with hexane/diethyl ether (1:1, v/v). The radioactiveproduct in the upper phase was concentrated to 1-2 ml by rotary evaporation, and further purified by preparative TLCon silica developed in diethyl ether/methanol (90:10). The band corresponding to 1-O-octadecyl[3H]glycol was scrapedand extracted twice with chloroform. The combined supernatants were evaporated to dryness, resuspended in absoluteethanol, and stored at 020 ЊC. Standard enzyme assay and lipid extraction. Membranes were obtained by disruption of stationary phase L. donovani
promastigotes (harvested at a density of 3-5 1 107 cells/ml) by nitrogen cavitation at 1500 psi for 25 minutes at 4ЊC(8). Large organelles and debris were removed by centrifugation at 10,000 1 g and the microsomal membranes werepelleted by centrifugation at 100,000 1 g. Cleavage activity was assayed by measuring the formation of [3H]ethyleneglycol from 1-O-octadecyl[3H]glycol cleavage. Standard incubation mixtures (200 ml) contained: membranes (1.5-2.0mg protein), 50 mM MES buffer, pH 7.0 (0.1 M Tris/HCl, 0.05 M MES, 0.05 M acetic acid, 0.25 M sucrose, 0.1 MKCl, 1 mM EDTA, 1 mM EGTA, 1mg/ml leupeptin, 0.1 mM TLCK), 5 mM (NH4)2SO4 containing 20 U/ml catalase,1.0 mM NADPH, 0.1 mM DTT, and 0.01-0.02 mCi 1-O-octadecyl[3H]glycol. The incubation mixture was incubatedat 35ЊC for 3.5 hours and was terminated by addition of 0.75 ml of chloroform/ methanol (1:2). Radioactivity in theaqueous phase was measured by scintillation counting. Analytical methods. Protein concentration was determined using the method of bicinchoninic acid microassay (9),
bovine serum albumin was used for a standard. In vitro analysis of alkylglycerol cleavage activity. In a previous report by Kaufman et al.
(7), an assay was developed for assaying alkylglycerol cleavage in a rat liver system usingalkylglycol as a substrate. Based on this assay, a cell lysate from L. donovani was examinedfor the capability of hydrolyzing 1-O-octadecyl[3H]glycol. The in vitro cleavage of the lipidsubstrate to form [3H]ethylene glycol was linear with time up to 1 h (Fig. 1A) and was protein
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Effects of Cofactors on Alkylglycol Cleavage Activity
Enzyme activity was assayed as described in Materials and Methods.
Background cpm (boiled enzyme control) was subtracted from the valuesshown.
concentration-dependent (Fig. 1B). Several possible reasons for why product formation wasnot linearly proportional to the amount of protein are inhibitory effects of increasing amountsof lipid (from the membranes) in the assay, consumption of substrate or cofactors, or increasedamounts of degradative enzymes. The pH optimum of the cleavage activity was 6.5-7 (Fig. 1C). In contrast to the mammalian system, pterin cofactors had no stimulatory effect oncleavage of the radioactive substrate whereas NADPH did stimulate Leishmania cleavageactivity (described below). Solubilization and partial purification of the alkylglycerol cleavage enzyme. The cleavage
activity was pelleted by centrifugation at 100,000 1 g, indicative of a membrane-boundenzyme. To solubilize the enzyme, this pellet was incubated with CHAPS at a final concentra-tion of 2.0% for 30 min at 4ЊC, and then was diluted to a CHAPS concentration of 0.3% foran additional 2 h. After centrifugation at 100,000 1 g for 1 hour, approximately 80% of theenzymatic activity was recovered in the supernatant. The solubilized enzyme was subjectedto ion exchange chromatography on a column of QAE-cellulose pre-equilibrated with 0.1%CHAPS in 20 mM Tris-HCl, pH 7.5. The bulk of the enzyme bound to the anionic supportand was displaced by isocratic elution with 0.1 M Tris/HCl, pH 7.5. The fractions containingthe main peak of enzymatic activity were pooled, and the enrichment in specific activity wasjudged to be 10-fold. Cofactor requirements for alkylglycerol cleavage activity. The cofactor requirements of
the partially purified alkylglycerol cleavage enzyme were investigated. NADPH stimulatedmonooxygenase activity (Table I) with an apparent Km around 10 mM. NADH (1 mM) wasapproximately 50% as effective as NADPH in stimulating cleavage activity. Addition oftetrahydrobiopterin and/or pteridine reductase (PTR1) (10) had no effect on cleavage activityin the absence of NADPH. Moreover, the addition of related pteridine derivatives tetrahy-dropterin with or without PTR1, dimethyltetrahydropteridine and 5,6,7,8-tetrahydropteridineor the pteridine reductase inhibitor methotrexate (1 mM) failed to alter ether bond cleavage(data not shown). Glutathione also had no effect on the activity. Similar cofactor results wereobtained using a solubilized preparation of the enzyme which had not been fractionated onQAE cellulose. Collectively, these results were consistent with the presence of a NADPH-dependent alkylglycerol cleavage enzyme in L. donovani.Substrates of the alkylglycerol cleavage enzyme. Since the radioactive substrate for the
alkylglycerol cleavage assay was not a physiological substance, it was important to confirmthat the enzyme hydrolyzes a biologically-relevant substrate. 1-O-octadecylglycerol is presentin the parasite and is a shorter chain version of the alkylglycerol portion of LPG and GIPLs. When 1-O-octadecylglycerol was used as the substrate in the in vitro assay, the cleavage of
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Inhibition of 1-O-octadecyl[3H]glycol cleavage by 1-O-octadecylglycerol. Unlabeled 1-O-octadecylglycerol
using the designated concentrations was added to the standard enzyme assay and the amount of [3H]ethylene glycolwas measured.
the ether bond was also observed and the aqueous-soluble product was identified as glycerolby paper chromatography. 1-O-octadecylglycerol was also examined for inhibition of cleavageof the 1-O-[3H]octadecylglycol analog in the alkylglycerol cleavage assay using QAE-cellulose-purified enzyme. Increasing the concentration of unlabeled 1-O-octadecylglycerol resulted ina decrease in cleavage of radiolabeled substrate (Fig. 2). At 1 mM of 1-O-octadecylglycerol,more than 95% of the radiolabeled glycol substrate remained uncleaved. These results indicatedthat the same alkylglycerol cleavage enzyme that catalyzed the cleavage of the alkylglycolwas also responsible for the degradation of alkylglycerol in L. donovani.
Tietz et al. (3) first reported the finding of a rat liver microsomal enzyme that could
cleave 1-O-alkylglycerol. Similar to phenylalanine, tyrosine and tryptophan hydroxylases, thismammalian enzyme had a strict requirement for tetrahydrobiopterin and molecular oxygen. Addition of pteridine reductase, which functions to reduce the ''oxidized pteridine'' intermedi-ate to the tetrahydro level, could stimulate enzyme activity. The cofactor-dependence of theliver glyceryl ether monooxygenase was later shown to require one of several different tetrahy-dropterins. The most effective pterins were the unconjugated tetrahydropterins rather thanconjugated derivatives, such as tetrahydrofolate. Unlike the rat liver enzyme system, ourresults indicated a major difference in Leishmania with respect to the requirement of reducingcofactors. NADPH was found to be the necessary cofactor for ether-cleaving activity. Neithertetrahydrobiopterin or related pteridines was involved in the reaction. Glyceryl ether monooxy-genase activity has also been investigated in Tetrahymena pyriformis. Similar to the Leishmaniaenzyme, cell-free preparations of T. pyriformis required NADPH to cleave the ether bond (11).
We observed other differences between the rat liver and Leishmania alkylglycerol cleaving
enzymes. Glutathione was necessary for expression of full enzyme activity in rat liver micro-somes (12). Since N-ethylmaleimide inhibits the rat liver enzyme activity, it was believed thatthe role of glutathione was probably to maintain the sulfhydryl groups of the enzyme. Incontrast, omission of glutathione in the Leishmania system had no effect on the enzymeactivity. The pH and temperature optimum of the L. donovani enzyme were also differentfrom those of the rat liver monooxygenase. All these results indicated that a distinct enzymein L. donovani was responsible for the cleavage of the ether bond.
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
The role of the monooxygenase in the parasite is unknown. Interestingly, the Leishmania
alkylglycerol cleavage enzyme was unable to hydrolyze the alkylglycerol anchor of purified,intact LPG or GIPLs. This observation suggests that alkylglycerol cleavage does not initiateLPG and GIPL turnover, but may be a later step in GPI lipid anchor metabolism. Alternatively,LPG and GIPLs may first need to be preprocessed or associated with another molecule priorto cleavage of the lipid anchor by the monooxygenase. It is also possible that the enzymedescribed in this study might not be involved in GPI lipid anchor turnover, but rather in themetabolism of alkylphospholipids, which are abundantly present in Leishmania (13). Thedifference in cofactor requirements for the mammalian and parasitic alkylglycerol monooxy-genase may represent a potential target for effective therapeutic protocols for the treatment ofthis disease if the enzyme is shown to be essential for parasitic metabolism. Full characterizationand purification of the Leishmania alkylglycerol monooxygenase should facilitate studies inthe generation of specific inhibitors of the enzyme.
The authors are indebted to Dr. Bakela Nare for the PTR1 protein and to Drs. Robert Lester and Charles Waechter
1. Turco, S. J., and Descoteaux, A. (1992) Annu. Rev. Microbiol. 46, 65 - 94. 2. McConville, M. J., and Ferguson, M. A. J. (1993) Biochem. J. 294, 305 - 324. 3. Tietz, A., Lindberg, M., and Kennedy, E. P. (1964) J. Biol. Chem. 239, 4081 - 4090. 4. Acterberg, V., and Gercken, G. (1987) Mol. Biochem. Parasitol. 23, 117 - 122. 5. King, D. L., and Turco, S. J. (1988) Mol. Biochem. Parasitol. 28, 285 - 294. 6. Orlandi, P. A., Jr., and Turco, S. J. (1987) J. Biol. Chem. 262, 10384 - 10391. 7. Kaufman, S., Pollock, R. J., Summer, G. K., Das, A. K., and Hajra, A. K. (1990) Biochim. Biophys. Acta 1040,
8. Carver, M. A., and Turco, S. J. (1991) J. Biol. Chem. 266, 10974 - 10981. 9. Smith, P. K., Krohn, R. I., Hermanson, G. T., Mallia, A. K., Gartner, F. H., Provenzano, M. D., Fujimoto, E. K.,
Goeke, B. J., Olson, B. J., and Klenk, D. C. (1985) Anal. Biochem. 150, 76 - 85.
10. Bello, A. R., Nare, B., Freedman, D., Hardy, L., and Beverley, S. M. (1994) Proc. Natl. Acad. Sci. USA 91,
11. Kapoulas, V. M., Thompson, G. A., Jr., and Hanahan, D. J. (1969) Biochim. Biophys. Acta 176, 250 - 264. 12. Ishibashi, T., and Imai, Y. (1983) Eur. J. Biochem. 132, 23 - 27. 13. Wassef, M. K., Fioretti, T. B., and Dwyer, D. M. (1985) Lipids 20, 108 - 115.
democratização da terra”, como mostramos anteriormente, é necessárioDISCURSOS SOBRE O ESTRANGEIRISMO EM GRAMÁTICAS:democratizar a terra, o que já foi feito em países capitalistas como EUA,Japão etc. Democratizar a terra é torná-la fonte de emprego, justiça social,alimentos e renda. É também necessário democratizar o poder, pontonevrálgico de resistência e dominação histó
 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. Analysis of alkylglycerol cleavage activity in a cell-free system of L. donovani. Membranes (2 mg/ml)
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 1. Analysis of alkylglycerol cleavage activity in a cell-free system of L. donovani. Membranes (2 mg/ml)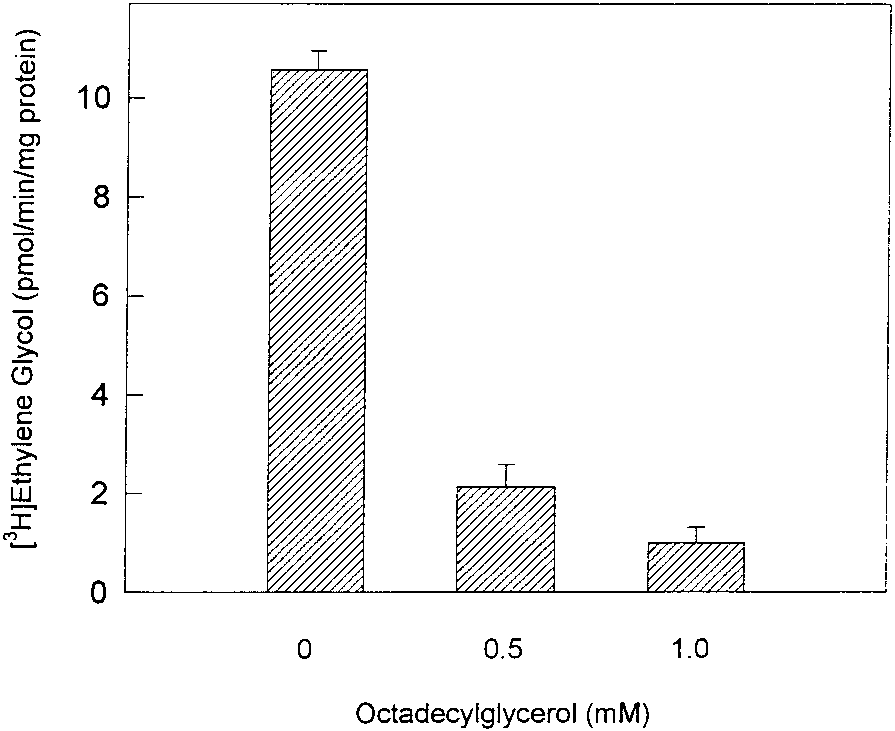 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Inhibition of 1-O-octadecyl[3H]glycol cleavage by 1-O-octadecylglycerol. Unlabeled 1-O-octadecylglycerol
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
FIG. 2. Inhibition of 1-O-octadecyl[3H]glycol cleavage by 1-O-octadecylglycerol. Unlabeled 1-O-octadecylglycerol