L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
485umb_tanaka.qxd
Naunyn-Schmiedeberg's Arch Pharmacol (2001) 364 : 538-550DOI 10.1007/s002100100485
Fumiko Yamaki. Momoko Kaga. Takahiro Horinouchi. Hikaru Tanaka. Katsuo Koike. Koki Shigenobu. Ligia Toro. Yoshio Tanaka
MaxiK channel-mediated relaxation of guinea-pig aorta following stimulation of IP receptor with beraprost via cyclic AMP-dependent and -independent mechanisms
Received: 12 March 2001 / Accepted: 24 August 2001 / Published online: 23 October 2001
Abstract The present study was aimed to elucidate the
did not affect beraprost-induced relaxation though it al-
cellular pathway(s) controlling vascular relaxation triggered
most totally inhibited the elevation of cyclic AMP con-
by stimulation of prostaglandin I2 (PGI2, IP) receptor tents induced by beraprost, suggesting the existence of anwith a stable PGI2 analog, beraprost. Beraprost caused a
additional mechanism that is cyclic AMP-independent.
concentration-dependent relaxation in de-endothelialized
Moreover, cholera toxin (CTX, 1 µg/ml for 6 h), which
guinea-pig aorta contracted with prostaglandin F2α (PGF2α).
activates the stimulatory G protein of adenylyl cyclase
Beraprost-induced relaxation was almost abolished in
(Gs), significantly suppressed PGF2α-induced contraction
high-KCl-contracted tissue, indicating a major role of K+
both in the absence and presence of SQ 22,536 (10-4 M).
conductances. In contrast to other PGI2 analogs (e.g. ci-
Iberiotoxin (10-7 M) was also capable of restoring the re-
caprost and iloprost), beraprost-induced relaxation was prac-
laxation induced by CTX. These findings suggest that
tically abolished by a selective voltage and Ca2+-activated
MaxiK channel plays a primary role in mediating smooth
K+ (MaxiK, BK) channel blocker Iberiotoxin (10-7 M) or
muscle relaxation following stimulation of IP receptor with
by tetraethylammonium (2×10-3 M). The relaxation in-
beraprost in guinea-pig aorta. Both cyclic AMP-depen-
duced by beraprost was not significantly affected by other
dent and -independent pathways contribute to the MaxiK
K+ channel blockers glibenclamide (10-6 M) or Ba2+ (10-5 M),
channel-mediated relaxation following IP receptor stimu-
but was slightly attenuated by 4-aminopyridine (10-4 M).
lation in this vascular tissue. Direct regulation of MaxiK
Beraprost increased intracellular cyclic AMP levels, sug-
channels by Gs may partly account for the cyclic AMP-in-
gesting a role for cyclic AMP-dependent pathways. A se-
lective inhibitor of cyclic AMP-specific phosphodiesterase,RO-20-1724 (10-4 M), significantly potentiated beraprost-
Keywords Adenosine 3':5'-cyclic monophosphate.
induced relaxation. Iberiotoxin (10-7 M) completely coun-
Beraprost. BK channel. Cholera toxin. Guinea-pig
teracted this potentiation. Moreover, tension decrement due
aorta. Iberiotoxin. MaxiK channel. Prostaglandin I2
to forskolin (3×10-7 M) or 8-bromo-cyclic AMP (10-2 M)
receptor. SQ 22,536. Stimulatory G protein of adenylyl
was thoroughly restored by Iberiotoxin (10-7 M), confirm-
ing a role for a cyclic AMP-dependent mechanism. How-ever, SQ 22,536 (10-4 M), an adenylyl cyclase inhibitor,
Prostaglandin I2 (PGI2, prostacyclin) is an arachidonic
F. Yamaki. M. Kaga. H. Tanaka. K. Shigenobu. Y. Tanaka (✉)
acid metabolite of the cyclooxygenase pathway, which is
Department of Pharmacology, Toho University School of Pharmaceutical Sciences,
synthesized mainly in vascular endothelial cells but also
Miyama 2-2-1, Funabashi-City, Chiba 274-8510, Japan
in vascular smooth muscle cells (Leffler 1997; Moncada
et al. 1977; Narumiya et al. 1999). Although PGI2 itself
Tel.: +81-47-4722099, Fax: +81-47-4722113
potently inhibits vascular tone as well as platelet aggrega-
tion, a number of stable PGI2 analogues have been devel-
oped for clinical use since the half-life of PGI2 is less than
Toho University School of Pharmaceutical Sciences,
Miyama 2-2-1, Funabashi-City, Chiba 274-8510, Japan
logues (iloprost, cicaprost, beraprost) inhibit vascular
contractions in guinea-pig aorta (Clapp et al. 1998; Ozaki
Departments of Anesthesiology and Molecular
et al. 1996), rat mesenteric and tail arteries (Adeagbo and
and Medical Pharmacology, and Brain Research Institute, UCLA School of Medicine, Los Angeles, CA 90095, USA
Malik 1990; Schubert et al. 1997), and dog arteries (cere-
bral, carotid, mesenteric, renal, femoral arteries; Akiba et
ing cyclic AMP-independent MaxiK channel activation
al. 1986; Siegel et al. 1989). Moreover, PGI2 analogues
and muscle relaxation? (4) Does stimulatory G protein
seem to be beneficial in the treatment of circulatory disor-
of adenylyl cyclase (Gs) produce MaxiK channel-medi-
ders especially for pulmonary hypertension (Wise and
ated vascular relaxation independent of cyclic AMP? In
Jones 1996). Thus, uncovering the mechanisms by which
the present study, we addressed these questions using
PGI2 and its analogues relax vascular smooth muscle is
guinea-pig aorta and a different PGI2 analog, beraprost
relevant from both physiological and therapeutic points of
(TRK-100; Akiba et al. 1986; Murata et al. 1989). Our re-
sults demonstrate that in guinea-pig aorta, both cyclic
Increasing electrophysiological and pharmaco-mechan-
AMP-dependent and cyclic AMP-independent mecha-
ical evidence suggests that PGI2 analogues exhibit their
nisms mediate the IP agonist-induced relaxation via acti-
vasorelaxant actions through activation of plasma mem-
vation of MaxiK channels. The cyclic AMP-independent
brane K+ channels, which leads to membrane hyperpolar-
mechanism is predominant in this vessel and cholera toxin
ization. Recent studies have focused on the role of the
experiments suggest that it may be due to a direct action
large conductance Ca2+-activated K+ (MaxiK, BK) chan-
nel in the relaxant effects of PGI2 analogues, iloprost andcicaprost (Clapp et al. 1998; Dumas et al. 1997; Schubertet al. 1996, 1997). It is likely that activation of MaxiK
channels with the resultant membrane hyperpolarizationlimits sarcolemmal Ca2+ influx through L-type voltage-
Hartley guinea-pigs of either sex (Oriental Yeast, Tokyo, Japan)were housed under controlled conditions (temperature 21°-22°C,
gated Ca2+ channels, and thus, produces vascular relax-
relative air humidity 50±5%). Food and water were available ad li-
ation by decreasing cytoplasmic Ca2+ levels ([Ca2+]cyt)
bitum to all animals. This study was conducted in accordance with
(Nelson and Quayle 1995; Toro et al. 1998). Consistent
the Guideline for the Care and Use of Laboratory Animals adopted
with the observations in pharmaco-mechanical studies, the
by the Committee on the Care and Use of Laboratory Animals ofToho University School of Pharmaceutical Sciences (accredited by
PGI2 receptor (IP receptor) agonist iloprost has been
the Ministry of Education, Science, Sports and Culture, Japan).
shown to increase MaxiK channel current in isolated arte-rial cells (Schubert et al. 1996, 1997). On the other hand,
Preparation of thoracic aortic rings. A section of the thoracic
in some blood vessels like rat cerebral (Lombard et al.
aorta between aortic arch and diaphragm was removed and placedin normal Tyrode's solution (mM): NaCl, 158.3; KCl, 4.0; CaCl
1999), pulmonary (Dumas et al. 1997) and tail arteries
(Schubert et al. 1997), and rabbit coronary vessels (Jack-
2, 1.05; NaH2PO4, 0.42; NaHCO3, 10.0 and glucose, 5.6.
The aorta was cleaned of loosely adhering fat and connective tis-
son et al. 1993), the ATP-sensitive K+ (KATP) channel
sue. To remove the endothelium, a thread moistened with normal
seems to also play a substantial role in the vascular relax-
Tyrode's solution was inserted into the internal side of the blood
vessel. The internal surface was gently rubbed by rolling the thread
using fingers on the palm covered with a rubber glove. The aorta
tion of K+ channel subtypes to vascular relaxations due to
was cut into ring segments about 2 mm in length.
PGI2 analogues may vary depending on the vascular bedemployed, and on the relative expression and/or colocal-
Measurement of tension changes. The ring segments were mounted
ization of different proteins in each vascular bed.
using stainless steel hooks (outer diameter, 200 µm) under the op-timal resting tension of 1.0 g (Tanaka et al. 2000) in a 5-ml organ
PGI2 and its analogs elevate intracellular adenosine
bath (UC-5; UFER Medical Instrument, Kyoto, Japan) containing
3':5'-cyclic monophosphate (cyclic AMP) levels due to
normal Tyrode's solution. Tension changes of the muscle prepara-
activation of adenylyl cyclase via the specific cell surface
tion were isometrically recorded with a force-displacement trans-
IP receptor-coupled guanine nucleotide regulatory pro-
ducer (T7-8-240; Orientec, Tokyo, Japan) connected to a minipoly-graph (RM-6100; Nihon Kohden, Tokyo, Japan or Signal Condi-
tein, Gs (Leffler 1997; Narumiya et al. 1999; Wise and
tioner: Model MSC-2; Labo Support, Suita-City, Japan). Ring prepa-
Jones 1996). Concomitantly in rat tail artery, it is thought
rations were equilibrated for 60-90 min prior contraction with iso-
that channel phosphorylation by cyclic AMP-dependent
tonic high-KCl (80 mM) Tyrode's solution (mM): NaCl, 82.3;
protein kinase (PKA) is responsible for the MaxiK chan-
KCl, 80.0; CaCl2, 2.0; MgCl2, 1.05; NaH2PO4, 0.42; NaHCO3,
nel-mediated vascular relaxation due to IP receptor stimu-
10.0 and glucose, 5.6. After washing out, the absence of endothe-lial cells was confirmed by the lack of relaxation induced by
lation by iloprost (Schubert et al. 1996, 1997). In contrast,
acetylcholine (ACh; 10-5 M), and experiments were started after a
in guinea-pig aorta, cyclic AMP-independent pathways
30-min equilibration period. Normal Tyrode's solution was contin-
were recently proposed to underlie the relaxation induced
uously gassed with 95% O2 - 5% CO2, and kept at 36.5±0.5°C
2 analogues, iloprost and cicaprost (Clapp et al.
1998; Turcato and Clapp 1999). Although these mecha-
Relaxant effects of test compounds on the contractions induced by
nistic differences may be attributed to the nature of the
prostaglandin F2α or high-KCl. To investigate the relaxation in-
vascular beds or species, there are several questions that
duced by test compounds beraprost, cromakalim, forskolin and
remain unanswered: (1) Do cyclic AMP-dependent and membrane-permeant cyclic nucleotides, aorta was contracted with
-independent mechanisms co-regulate MaxiK channel and
2α (PGF2α; 10-5 M) or high-KCl (80 mM). After the
muscle contractions reached a steady-state level, test compounds
vascular smooth muscle relaxation mediated via IP recep-
were applied cumulatively to the bath medium. Concentration-re-
tor occupancy in the same vessel (i.e. guinea-pig aorta)?
sponse relationships were obtained by measuring the relaxant ef-
(2) Is MaxiK channel activation involved in the IP recep-
tor-triggered relaxation mediated via cyclic AMP-inde-
Effects of K+ channel blockers, RO-20-1724, SQ 22,536 and ODQ.
pendent pathways? (3) What are the mechanisms underly-
Possible involvement of the activation of K+ channels in drug-in-
duced vasorelaxation was examined in the absence and presence of
penta [b] benzofuran-5-butyrate (beraprost sodium) was kindly do-
several K+ channel blockers. K+ channel blockers used in the pre-
nated by Toray Industries (Kamakura-City, Kanagawa, Japan).
sent study were: Iberiotoxin (10-7 M) and tetraethylammonium
Cromakalim was a kind gift from Nissan Chemical Industries
(TEA; 2×10-3 M) as MaxiK channel blockers; glibenclamide (Tokyo, Japan). Other chemicals were: prostaglandin F2α (PGF2α),
(10-6 M) as KATP channel blocker; Ba2+ (10-5 M) as inward recti-
7-[3-[[2-[(phenylamino)carbonyl]hydrazino]methyl]-7-oxabicyclo
fier K+ channel blocker; and 4-aminopyridine (4-AP; 10-4 M) as
[2.2.1]hept-2-yl]-, [1S-[1α,2α(Z),3α,4α]]-5-heptenoic acid (SQ
delayed rectifier K+ (Kv) channel blocker. Iberiotoxin, TEA, Ba2+
29,548) (Cayman Chemical, Ann Arbor, Mich., USA); l-noradren-
and 4-AP were added to the bath 20 min before starting cumulative
aline bitartrate (NA), forskolin, papaverine hydrochloride (Wako
application of test compounds. Glibenclamide was applied to the
Pure Chemical Industries, Osaka, Japan); acetylcholine chloride
bath solution 20 min before application of PGF2α. For forskolin-
(ACh; Daiichi, Tokyo, Japan); Iberiotoxin (Peptide Institute, Minoh-
and 8-bromo-cyclic AMP-induced relaxation, Iberiotoxin (10-7 M)
Shi, Osaka, Japan); glibenclamide (Sigma, St. Louis, Mo., USA);
was applied to the bath medium after the relaxant responses had
8-bromo-cyclic AMP, dibutyryl cyclic AMP (BIOLOG Life Sci-
reached a steady-state level; in this case, the reversion of relax-
ence Institute, Bremen, Germany); 9-(tetrahydro-2-furanyl)-9H-
purin-6-amine (SQ 22,536) (Research Biochemicals International,
RO-20-1724 (10-4 M; 4-(butoxy-4-methoxybenzyl)-2-imidazo-
Natick, Mass., USA); 4-(butoxy-4-methoxybenzyl)-2-imidazolidi-
lidinone), a selective inhibitor of cyclic AMP-specific phosphodi-
none (RO-20-1724), 1H-[1,2,4]oxadiazolo[4,3a]quinoxaline-1-one
esterase (soluble cyclic GMP-insensitive type IV PDE; Katano and
(ODQ) (Biomol Research Laboratories, Plymouth Meeting, Pa.,
Endoh 1990), was added to the bath solution 20 min before stimu-
USA); nitroglycerin (Nihon Kayaku, Tokyo, Japan); cholera toxin
lation with PGF2α (10-5 M) or after beraprost (10-7 M)-induced re-
(CTX; List Biological Laboratories, Campbell, Calif., USA). All
laxation reached a steady-state level.
other reagents used were commercially available and of reagent
SQ 22,536 (10-4 M; 9-(tetrahydro-2'-furyl)adenine), an adenylyl
cyclase inhibitor, was added to the bath solution 20 min before cu-
PGF2α, forskolin and RO-20-1724 were dissolved in pure
mulative application of beraprost. ODQ (10-5 M; 1H-[1,2,4]oxa-
ethanol as a stock solution at 10-2 M. Cromakalim was dissolved in
diazolo[4,3a]quinoxaline-1-one), an inhibitor of soluble guanylyl
70% ethanol at 10-2 M. Glibenclamide was dissolved in pure di-
cyclase, was applied to the bath solution when the relaxation due
methyl sulfoxide (DMSO) at 10-3 M. SQ 29,548 was dissolved in
to beraprost (10-5 M) or nitroglycerin (10-5 M) reached a steady-
100% ethanol at 10-2 M. ODQ was dissolved in 100% DMSO at
10-2 M. Dilutions were performed in water. Final ethanol andDMSO concentrations in the bath medium did not exceed 1% and
Determination of tissue cyclic AMP contents. Thoracic aortas were
0.1%, respectively, which did not affect the vascular responses. All
cut into segments about 15 mm in length and cleaned of loosely
drugs are expressed in molar concentrations in bathing solution.
adhering fat and connective tissue. Endothelium was removed asdescribed for contraction studies. Each aorta was opened along the
Analysis and statistics. The percentage of relaxation was calcu-
longitudinal axis of the artery to prepare a flat sheet. Each sheet
lated by considering the maximum tension level obtained just be-
was incubated in organ bath containing normal Tyrode's solution
fore addition of vasorelaxants as 0% relaxation, and the full recov-
(25 ml) which was continuously gassed with 95% O2 - 5% CO2
ery to the basal tension before application of PGF2α (10-5 M), NA
and maintained at 36.5±0.5°C (pH=7.35). After 60-min incuba-
(3×10-6 M) or high-KCl (80 mM) as 100% relaxation. Data were
tion, artery segments were exposed to beraprost (10-5 M) or
plotted as a function of drug concentrations and fitted to the equa-
forskolin (10-5 M) for 20 min. When treated with SQ 22,536, this
adenylyl cyclase inhibitor (10-4 M) was applied to the bath me-
dium 20 min before stimulation with beraprost. At the end of the
protocol, tissues were rapidly frozen in liquid N2 to terminate the
where E is the % relaxant response at a given drug concentration,
reaction. The arteries were homogenized in 6% trichloroacetic acid
(TCA) solution and then centrifuged at 3000 rpm for 15 min at
max is the maximum relaxant response, A is the concentration of
5°C. The supernatant fraction and the tissue pellets were used for
H is the slope function and EC50 is the effective drug
concentration that produces a 50% response. Curve fitting was per-
the measurement of cyclic AMP and protein contents, respectively.
formed using GraphPad Prism (version 2.01; GraphPad Software,
The cyclic AMP in the supernatant was extracted with water-
saturated ether for four times to remove TCA and then lyophilized.
Cyclic AMP contents were measured using an enzyme-immunoas-
50 (the drug concentration required to induce
50% inhibition) were also determined from the concentration-re-
say system (Amersham Pharmacia Biotech UK, Buckinghamshire,
sponse relationship of individual experiments and expressed as
UK). Tissue pellets were dissolved in 2 ml NaOH for protein de-
termination (Lowry et al. 1951). The cyclic AMP contents were
50 (negative logarithm of IC50) for statistical analysis.
Data are presented as means ± SEM and n refers to the number
expressed as picomoles per milligram of sample protein. In these
of experiments. The significance of the difference between mean
measurements, vascular tissues were not exposed to phosphodi-
values was evaluated with GraphPad Prism by paired or unpaired
esterase inhibitors since the inhibitors could elevate cyclic AMP to
t-test, unpaired t-test with Welch's correction if necessary, and
extremely unphysiologically high levels, and as a consequence
one-way analysis of variance (ANOVA) followed by Tukey's mul-
could elevate cyclic GMP levels, both of which may complicate
tiple comparison test. A P-value less than 0.05 was considered sta-
the interpretation of data (Turcato and Clapp 1999). Inhibitory effects of cholera toxin on PGF2α-induced contraction.
Aortic tissues were pre-contracted with PGF2α (10-5 M) and then
were incubated with cholera toxin (CTX; 1 µg/ml) in the continued
presence of PGF2α for 6 h. During the incubation with CTX, bath
solution was exchanged after 3 h with a fresh solution containing
Beraprost induces relaxation of guinea-pig aorta mainly
PGF2α (10-5 M) plus CTX (1 µg/ml). In a separate series of exper-
iments, aortic tissues were first contracted with PGF2α (10-5 M)
and washed out with fresh normal Tyrode's solution. After themuscle tension returned to the basal level, the tissue was incubated
First, we determined whether activation of K+ channels is
with CTX (1 µg/ml) for 6 h with a fresh solution exchange after involved in the relaxation induced by the IP receptor ago-3 h during the incubation. After 6 h, the muscle tissue was again
nist beraprost in guinea-pig aorta. This was investigated
by comparing the beraprost-induced relaxation in muscles
Sodium (±)-(1R,2R,3aS,8bS)-2,3,3a,8b-tetrahydro-2-hy-
contracted with PGF2α and high-KCl, which was used to
droxy-1-[(E)-(3S)-3-hydroxy-4-methyl-1-octen-6-ynyl]-1H-cyclo-
abolish the driving force for K+ efflux and the subsequent
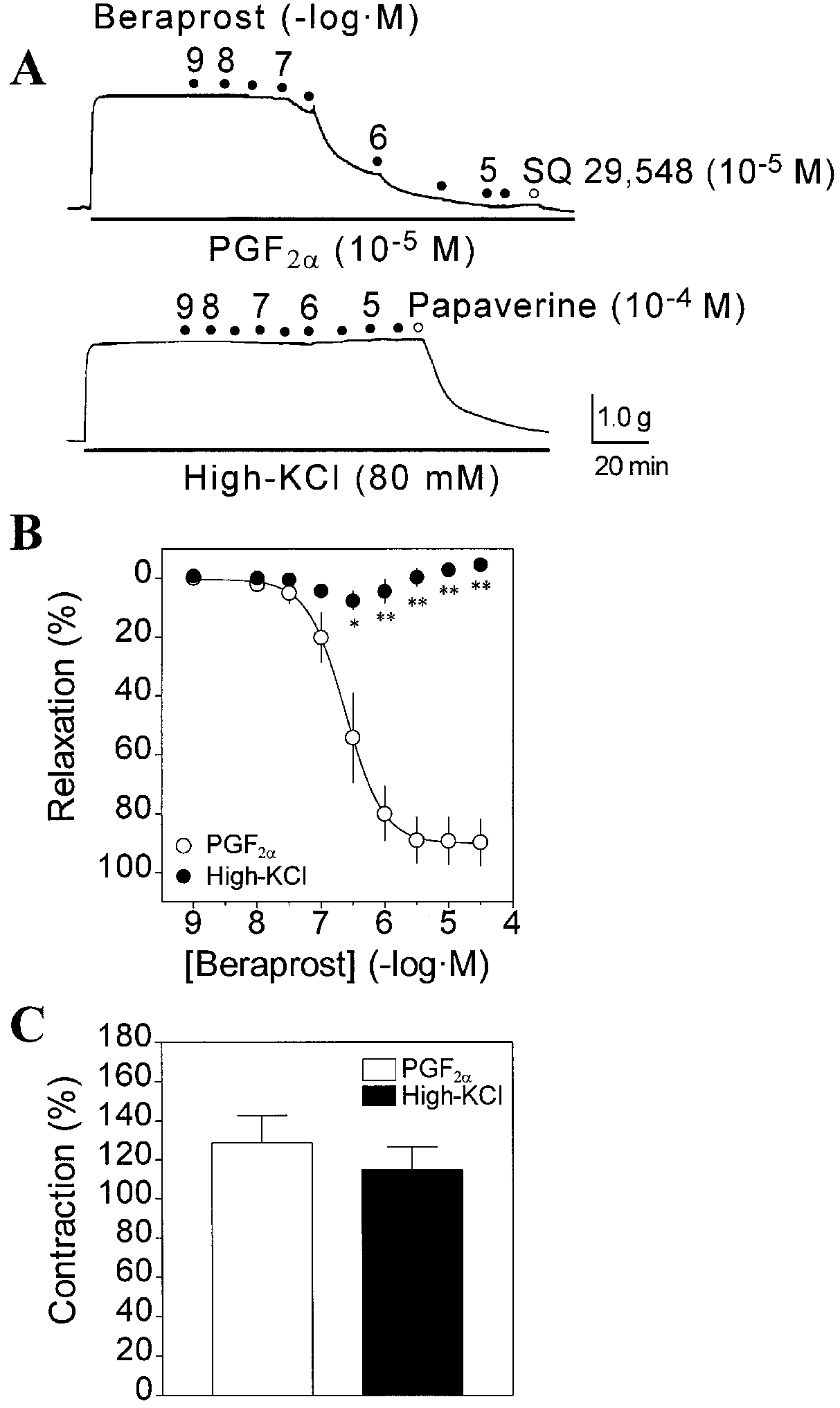
At concentration ranges over 10-6 M, a small contractioninstead of relaxation was attained by beraprost (Fig. 1B). Note that PGF2α and high-KCl induced similar contrac-tion levels (Fig. 1C), indicating that the lack of beraprost-induced relaxation with high-KCl is not due to an excesscontractile response with KCl. pIC50 values for beraprostagainst the contractions induced by PGF2α (10-5 M) andhigh-KCl (80 mM), and the maximum relaxant responsesare shown in Table 1. These observations strongly indi-cate that the relaxation of guinea-pig aorta in response toberaprost is mainly mediated via an activation of plas-malemmal K+ channels.
MaxiK channels are key contributors of beraprost-induced relaxation
MaxiK channel activation partially contributes (~40%-50%) to the relaxation induced by the IP agonists iloprostand cicaprost (Clapp et al. 1998). However, it is notknown whether MaxiK channels are involved in bera-prost-induced relaxation and to what extent. As shown inFig. 2A,B, beraprost-induced relaxation was concentra-tion-dependent and was strongly diminished in the prepa-rations treated with the MaxiK channel blocker, Ibe-riotoxin (10-7 M). Similarly, tetraethylammonium TEA (2×10-3 M), which blocks MaxiK channels at this concen-tration, strongly inhibited beraprost-induced relaxation (Fig. 2C). Note that MaxiK channels contribute by ~80%to the beraprost-induced relaxation as assessed by bothMaxiK channel blockers (Iberiotoxin and TEA; Fig. 2B,C;Table 1). These observations strongly indicate that activa-tion of MaxiK channel plays a major role in the relaxationinduced by beraprost in guinea-pig aorta, and that be-
Fig. 1A-C High-KCl suggests a primary role of K+ channels in beraprost-induced relaxation of guinea-pig aorta. A Typical me-
raprost is a better MaxiK channel activator than iloprost
chanical traces showing the relaxant effect of beraprost on the con-
traction induced by PGF2α (10-5 M; upper trace) or high-KCl
(80 mM; lower trace). Numbers are the negative logarithm of be-raprost concentration. SQ 29,548, an antagonist of PGF2α, was
added at the end of the experiment and produced near 100% relax-
ation. Papaverine was used to test the ability of the muscle to relax. B Concentration-response relationships for the relaxant effect of beraprost in rings precontracted with PGF2α (❍) or high-KCl (●).
The effect of glibenclamide on beraprost-induced relax-
Percentage of vascular relaxation is calculated with respect tobasal tension (100% relaxation) and PGF
ation was examined to determine whether the activation
(80 mM) steady-state contractile responses (0% relaxation). Data
of ATP-sensitive K+ (KATP) channel significantly con-
are means ± SEM of 4-6 experiments. *P<0.05, **P<0.01 signif-
tributes to the relaxant response as is the case of iloprost
icant differences between two groups. C Contractions of guinea-
in rat arteries (Dumas et al. 1997; Schubert et al. 1997).
pig aorta in response to PGF2α (10-5 M) and high-KCl (80 mM).
Glibenclamide (10-6 M) was not able to prevent be-
The contractions are normalized with respect to high-KCl (80 mM)-induced muscle tension obtained at the beginning of experiments
raprost-induced relaxation in guinea-pig aorta (Fig. 3A;Table 2). As control, when cromakalim, a KATP channelopener, was used to cause relaxation, glibenclamide
membrane hyperpolarization. Figure 1A shows represen-
(10-6 M) almost abolished its relaxant effect (Fig. 3B).
tative traces for the relaxant effect of beraprost on the
These findings rule out the role of KATP channel as a
mechanism by which beraprost causes relaxation of
lated guinea-pig aorta. Beraprost (3×10-8-3×10-5 M) clearly
elicited a concentration-dependent relaxation in the mus-
In the next series of experiments, we examined the ef-
fects of Ba2+ (an inward rectifier K+ channel blocker) and
contracted with noradrenaline (3×10-6 M; not shown). On
4-AP (a voltage-dependent K+ (Kv) channel blocker) to
the other hand, beraprost-induced relaxation was strongly
determine whether the activation of these K+ channels
diminished in vessels contracted with high-KCl (80 mM).
contributes to beraprost-induced relaxation. Ba2+ (10-5 M)
Table 1 Effects of high-KCl and MaxiK channel blockers on the
beraprost; also represented as % change with respect to high-KCl
relaxation induced by beraprost in guinea-pig aorta. Tension (g) is
(80 mM)-induced contraction obtained at the beginning of experi-
the muscle tension developed by PGF2α, high-KCl and PGF2α in
ments. pIC50 and Emax values refer to the relaxation induced by
the absence or presence of MaxiK channel blockers (Iberiotoxin;
3×10-5 M beraprost. Results are represented as means ± SEM of n
tetraethylammonium, TEA) just before cumulative application of
an=5bResponse obtained by 3×10-7 M beraprost**P<0.01 significant differences between two groups
Fig. 2A-C MaxiK channel blockers prevent the relaxation induced
concentrations, suggesting a minor role of Kv channels in
by beraprost in guinea-pig aorta. A Typical mechanical traces
the beraprost-induced relaxation of guinea-pig aorta.
showing the relaxant effect of beraprost on the contraction induced
Higher concentrations of Ba2+ and 4-AP were difficult to
by PGF2α (10-5 M) in the absence (left trace) and presence (right
assess in this blood vessel since both agents caused an in-
trace) of Iberiotoxin (IbTX, 10-7 M). Numbers are the negative log-
crease in basal tension (not shown).
arithm of beraprost concentration. Addition of SQ 29,548 (10-5 M), an antagonist of PGF2α, produced almost 100% relaxation. B, C
Concentration-response relationships for beraprost-induced vascu- lar relaxation in the absence (❍, control) and presence (●) of Iberiotoxin (10-7 M; B) and TEA (2×10-3 M; C). Vascular relax-
An inhibitor of cyclic AMP-specific phosphodiesterase,
ation was calculated with respect to basal tension (100% relax-
RO-20-1724, reveals a cyclic AMP-dependent
ation) and steady-state contractile responses to PGF2α (10-5 M) or
PGF2α plus Iberiotoxin or TEA (0% relaxation). Note that Iberio-
toxin (10-7 M) and TEA (2×10-3 M) further enhanced muscle ten-sion due to PGF2α from 1.88±0.09 g to 1.95±0.10 g (n=6 for each,
P<0.05), and from 2.48±0.15 g to 2.73±0.18 g (n=5 for each,
Cyclic AMP has been thought to be the key intracellular
P>0.05), respectively. Data are means ± SEM of 5-6 experiments.
messenger mediating vascular smooth muscle relaxation
*P<0.05, **P<0.01 significant differences between two groups
after stimulation of IP receptors (Kukovetz et al. 1979;Miller et al. 1979; Ozaki et al. 1996). However, in guinea-
did not significantly affect beraprost-induced relaxation
pig aorta an inhibitor of adenylyl cyclase, SQ 22,536, did
(Fig. 3C; Table 2). However, 4-AP (10-4 M; Fig. 3D; Table 2)
not prevent relaxation by the IP agonist iloprost, suggest-
had a modest but significant inhibitory effect at higher
ing that iloprost-induced relaxation is exclusively medi-
Fig. 3A-D 4-Aminopyridine (4-AP) but not glibenclamide or
(n=4 for each, P>0.05), and 2.10±0.23 g to 2.21±0.24 g (n=6,
Ba2+ (10-5 M) affect beraprost-induced relaxation in guinea-pig
P<0.05), respectively. Data are means ± SEM of 4-7 experiments.
aorta. A,C,D Concentration-response relationships for the relax-
*P<0.05 significant differences between two groups. B Concentra-
ation induced by beraprost in the absence (❍, control) and pres-
tion-response relationships for the relaxation induced by cro-
ence (●) of glibenclamide (10-6 M; A), Ba2+ (10-5 M; C) and 4-AP
makalim in the absence (❍, control) and presence (●) of gliben-
(10-4 M; D). Vascular relaxation was calculated with respect to
clamide (10-6 M). Vascular relaxation is expressed as percent inhi-
basal tension (100% relaxation) and steady-state contractile re-
bition against the tension elevation just before addition of cro-
sponses to PGF2α (10-5 M) or PGF2α plus Ba2+ or 4-AP (0% relax-
makalim. Data are means ± SEM of four experiments. *P<0.05,
ation). Note that Ba2+ (10-5 M) and 4-AP (10-4 M) changed the
**P<0.01 significant differences between two groups
muscle tension due to PGF2α from 2.11±0.17 g to 2.18±0.18 g
Table 2 Effects of K+ channel blockers, RO-20-1724 and SQ
22,536 (adenylyl cyclase inhibitor) just before cumulative admin-
22,536 on the relaxation induced by beraprost in guinea-pig aorta
istration of beraprost; also represented as % change of high-KCl
contracted with PGF2α (10-5 M). Tension (g) is the muscle tension
(80 mM)-induced contraction obtained at the beginning of experi-
development due to PGF2α (10-5 M; control) or PGF2α (10-5 M)
ments. pIC50 and Emax values refer to the relaxant effects of 3×10-5
plus K+ channel blockers (glibenclamide, Ba2+, 4-aminopyridine),
M beraprost. Results are represented as means ± SEM of n number
RO-20-1724 (cAMP-specific phosphodiesterase inhibitor) or SQ
*P<0.05 significant differences between two groups
ated by cyclic AMP-independent mechanisms (Turcato
slightly inhibited by the K+ channel blocker TEA (Clapp
and Clapp 1999). In line with this view, relaxation by
et al. 1998), raising the possibility (but not demonstrating)
forskolin, an activator of adenylyl cyclase, was only
that IP-agonists activate MaxiK channels solely by cyclic
Fig. 4B). Note that muscle tension decrement due to RO-20-1724 (10-4 M) alone was only 3.9±1.2% (n=3). These mechanical studies with RO-20-1724 indicate thatberaprost elevates intracellular cyclic AMP contents, andprovide direct evidence that cyclic AMP-dependentmechanisms also participate in beraprost-induced relax-ation via opening of MaxiK channels in this blood vessel.
Forskolin and 8-bromo-cyclic AMP mimic the beraprost-induced relaxation via activation of MaxiK channels
In the next series of experiments, we bypassed IP-receptorstimulation and determined whether cyclic AMP-mediatedrelaxation can be attributed to the activation of MaxiKchannels using the specific blocker Iberiotoxin. For thispurpose, we used two agents as vasorelaxants: forskolin, adirect activator of adenylyl cyclase, and 8-bromo-cyclicAMP, a membrane permeant cyclic AMP analogue. Fig-ure 5A shows the effects of pretreatment with Iberiotoxinon the relaxation induced by forskolin. Forskolin (10-9-10-5 M) fully relaxed PGF2α (10-5 M)-contracted musclein a concentration-dependent fashion. As previously ob-served for TEA (Clapp et al. 1998), forskolin-induced re-
Fig. 4 A, B Potentiation of beraprost-induced relaxation by a cyclic
laxation was not significantly affected by pretreatment
AMP-specific phosphodiesterase inhibitor, RO-20-1724: inhibi-
with Iberiotoxin (10-7 M). However, forskolin-induced re-
tion by a MaxiK channel blocker. A Concentration-response rela-
laxation was substantially diminished in the muscle con-
tionships for beraprost-induced relaxation in the absence (❍, con-trol) and presence (●) of RO-20-1724 (10-4 M). Vascular relax-
tracted with high-KCl (80 mM). pIC50 value of forskolin
ation is expressed as percent inhibition against the tension eleva-
against high-KCl-induced contraction (5.72±0.28, n=6)
tion due to PGF2α (10-5 M) just before addition of beraprost. Data
was significantly different from the values obtained with
are means ± SEM of six experiments. *P<0.05 significant differ-
PGF2α-induced contractions in the absence and presence
ences between two groups. B Iberiotoxin inhibits the RO-20-1724-
of Iberiotoxin (6.90±0.27, n=13 and 6.91±0.19, n=8;
induced potentiation of relaxation by beraprost. RO-20-1724 (10-4 M) was applied to the bath after beraprost (10-7 M)-induced
P<0.01, one-way ANOVA), indicating the participation of
relaxation reached a steady-state level. Iberiotoxin (10-7 M) was
K+ channels in the forskolin-induced relaxation. There-
administered after the relaxant response of beraprost was potenti-
fore, we decided to apply Iberiotoxin (10-7 M) during the
ated by RO-20-1724. RO-20-1724 (10-4 M) alone had a small re-
muscle relaxation with forskolin. In this case, the toxin
laxant effect. Data are means ± SEM of 3-4 experiments. Meanabsolute values of the tension development in response to PGF
completely counteracted the tension decrease caused by
(10-5 M) were 2.51±0.20 g (n=4; preparations used for the relax-
forskolin (3×10-7 M; Fig. 5B). Consistent with a role of
ation by beraprost) and 2.13±0.43 g (n=3; preparations used for the
MaxiK channels in forskolin-induced relaxation, pretreat-
relaxation by RO-20-1724). a)**P<0.01 significant differences
ment with TEA (2×10-3 M) slightly but significantly at-
from control (beraprost alone), b)**P<0.01 significant differencesfrom the response to beraprost (10-7 M) plus RO-20-1724 (10-4 M)
tenuated forskolin-induced relaxation at some concentra-tions (Fig. 5C), which was further confirmed by a signifi-cant decrease in the pIC50 value (control, 6.54±0.18, n=6
AMP-independent mechanisms in this blood vessel.
vs. TEA, 5.95±0.16, n=8; P<0.05).
Therefore, we examined the role of cyclic AMP and di-
To elucidate further the role of cyclic AMP in MaxiK
rectly tested the participation of MaxiK channels in the re-
channel-mediated vascular relaxation, the effect of Iberio-
laxation induced by beraprost in guinea-pig aorta. To this
toxin on the relaxation induced by 8-bromo-cyclic AMP
end, we used several pharmacological tests on beraprost-
was investigated. In this vascular preparation, 8-bromo-
induced relaxation using agonists and antagonists of cyclic
cyclic AMP elicited relaxation at concentration ranges
over 10-3 M, and the relaxation obtained by 10-2 M 8-bromo-
Figure 4A shows the effect of the cyclic AMP-specific
cyclic AMP was 61.0±6.9% (n=6; Fig. 5D). A similar
phosphodiesterase inhibitor RO-20-1724 on the relaxation
concentration-response relationship was also obtained for
induced by beraprost. In the presence of RO-20-1724
the relaxation induced by another membrane-permeant
(10-4 M), beraprost-induced relaxation was significantly
cyclic AMP analog, dibutyryl-cyclic AMP (pIC50, 2.76;
potentiated, as seen from the significant increases in both
Emax, 74.3% at 3×10-3 M, n=2 for each). In this case, ap-
the pIC50 value and the maximum relaxant response (see
plication of Iberiotoxin during relaxation was not neces-
also Table 2). Furthermore, this potentiation was com-
sary since pretreatment with Iberiotoxin (10-7 M) almost
pletely counteracted by addition of Iberiotoxin (10-7 M;
completely eliminated 8-bromo-cyclic AMP-elicited re-
Fig. 5A-E MaxiK channel blockers also inhibit the relaxations in-
laxation (Fig. 5D vs. Fig. 5A). As expected, when Iberio-
duced by forskolin and 8-bromo-cyclic AMP in guinea-pig aorta.
toxin (10-7 M) was applied during the muscle relaxation,
A,B Iberiotoxin inhibits forskolin-induced relaxation depending
it completely restored the tension previously decreased by
on the application sequence. Iberiotoxin (10-7 M) applied prior
10-2 M 8-bromo-cyclic AMP (Fig. 5E) as with the case of
forskolin treatment was unable to prevent forskolin-induced relax- ation (A). However, Iberiotoxin (10-7 M) applied during forskolin
forskolin (Fig. 5B). These results strongly suggest that
(3×10-7 M)-induced relaxation was able to fully prevent this relax-
cyclic AMP-mediated mechanisms contribute to the acti-
ation (B). Consistent with a K+ channel role, high-KCl (80 mM)
vation of MaxiK channel in guinea-pig aorta.
also prevented forskolin-induced relaxation (A). Vascular relax- ation is expressed as percent inhibition against the tension devel- opment due to PGF2α (10-5 M; ❍, control), PGF2α (10-5 M) plus
Iberiotoxin (10-7 M; ●), or high-KCl (80 mM; ■), just before ad-
Inhibition of cyclic AMP production with SQ 22,536
ministration of forskolin. Mean absolute values of the tension de-
does not affect beraprost-induced relaxation
velopment prior addition of forskolin were (A): 1.90±0.12 g (n= 13; control, 10-5 M PGF2α); 2.08±0.15 g (n=8; 10-5 M PGF2α plus
10-7 M Iberiotoxin); 2.02±0.13 g (n=6; 80 mM KCl). Data are
The mechanical studies using RO-20-1724 and cyclic AMP
means ± SEM of 5-13 experiments. A a)*P<0.05 significant dif-
elevating agents can lead to the hypothesis that stimula-
ference from control; b)*P<0.05, b)**P<0.01 significant difference
tion of IP receptor with beraprost elevates cyclic AMP
from the response in the presence of Iberiotoxin. B *P<0.05 sig- nificant difference from the response in the absence of Iberiotoxin.
contents, and thus, causes MaxiK channel-mediated relax-
C Inhibitory effect of TEA on forskolin-induced relaxation. TEA
ation. If the cyclic AMP-mediated pathway is one of the
(2×10-3 M) was applied prior forskolin treatment. Mean absolute
mechanisms by which beraprost elicits MaxiK channel-
values of the tension development just before cumulative addition
mediated relaxation, the inhibition of adenylyl cyclase
of forskolin are 1.93±0.28 g (n=6; ❍, control, 10-5 M PGF2α) and
should diminish vasorelaxation induced by this IP ago-
1.96±0.18 g (n=8; ●, 10-5 M PGF2α plus 2×10-3 M TEA). Data are
means ± SEM of 6-8 experiments. *P<0.05 significant differences
nist. Thus, we inhibited adenylyl cyclase and examined its
between two groups. D,E Iberiotoxin inhibits 8-bromo-cyclic
effect on beraprost-induced vascular relaxation. As shown
AMP-induced relaxation regardless of the addition sequence. Iberio-
in Fig. 6A and Table 2, SQ 22,536 (10-4 M), an inhibitor
toxin (10-7 M) was applied prior 8-bromo-cyclic AMP (8-Br-cAMP)
of adenylyl cyclase, did not significantly affect the relax-
(D) or during 8-bromo-cyclic AMP-induced relaxation (E). Mean absolute values of the tension development prior 8-bromo-cyclic
ant response to beraprost. These results are similar to those
AMP treatment are: 2.43±0.17 g (n=6; ❍, control, 10-5 M PGF2α)
obtained with iloprost (Turcato and Clapp 1999).
and 2.82±0.27 g (n=5; ●, 10-5 M PGF2α plus 10-7 M Iberiotoxin).
Figure 6B shows the increase in cyclic AMP content in
In E, the control experiment corresponds to the last addition of response to beraprost and its inhibition by SQ 22,536. Be-
8-bromo-cyclic AMP (10-2 M) of the control concentration-response curve in D, at which point Iberiotoxin (10-7 M) was added. Data
raprost (10-5 M) elevated cyclic AMP content from
are means ± SEM of 5-6 experiments. *P<0.05, **P<0.01 signif-
2.5±0.5 pmol per mg protein to 75.9±10.8 pmol per mg
protein (n=6 for each, P<0.01), producing a 30.4-fold rise. In contrast, cyclic AMP content was not significantly ele-vated due to the stimulation with beraprost (10-5 M) in thepresence of SQ 22,536 (10-4 M). SQ 22,536 (10-4 M) it-
able to relax the vascular muscle even when the adenylylcyclase activity is strongly inhibited by SQ 22,536 (Fig. 6C). Furthermore, Fig. 6C shows that the elevation of cyclicAMP content is much less by beraprost than by forskolinfor generating the same amount of muscle relaxation. Thepharmaco-mechanical studies with SQ 22,536 togetherwith the measurement of cyclic AMP content suggest thatrelaxation induced by beraprost is primarily produced viacyclic AMP-independent pathways. However, experi-ments performed with RO-20-1724 indicate that beraprostmay trigger a secondary cyclic AMP-mediated path-way(s) that produces relaxation and that this pathway ac-tivates MaxiK channels. This view is supported by thefact that Iberiotoxin inhibits forskolin- and 8-bromo-cyclic AMP-induced relaxations.
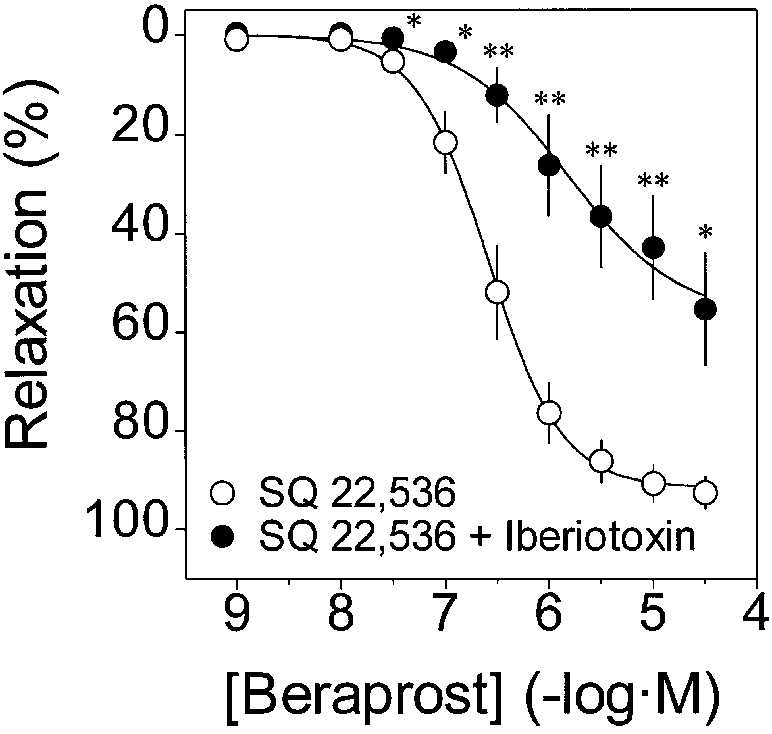
MaxiK channels participate in the cyclic AMP-independent component of beraprost-induced relaxation
Since the involvement of MaxiK channels in the cyclicAMP-independent component of IP agonist-induced re-laxation has not been tested experimentally, we directlyblocked channel activity with Iberiotoxin to determinetheir role. We now show that Iberiotoxin (10-7 M) signifi-cantly diminishes the beraprost-induced relaxant compo-nent in the presence of SQ 22,536 (10-4 M; Fig. 7). BothpIC50 (control, 6.50±0.14, n=7 vs. Iberiotoxin, 5.49±0.39,n=8, P<0.05) and the maximum response values (at 3×10-5 M beraprost; control, 92.6±3.4%, n=7 vs. Iberio-toxin, 50.1±10.4%, n=8, P<0.01) were significantly atten-uated by Iberiotoxin. Note that the maximum inhibition atequivalent beraprost values (>1×10-6 M) was ~80% in be-
Fig. 6A-C SQ 22,536 inhibits production of cyclic AMP but has negligible effect on beraprost-induced relaxation in guinea-pig
raprost-induced relaxations without SQ 22,536 (Fig. 2),
aorta. A Concentration-response relationships for beraprost-in-
whereas it was only ~50% when the adenylyl cyclase path-
duced relaxation in the absence and presence of the adenylyl cy-clase inhibitor SQ 22,536 (10-4 M). SQ 22,536 did not affect be-raprost-induced relaxation. Vascular relaxation is expressed as apercent inhibition against the muscle contraction induced byPGF2α (10-5 M). Data are means ± SEM of 3-4 experiments.
B Basal cyclic AMP content (control) was not affected by SQ 22,536 (10-4 M, SQ). Beraprost (10-5 M, Bera) increased tissue cyclic AMP content, which was inhibited by SQ 22,536. Data are means ± SEM of 5-6 experiments (in duplicates). **P<0.01 sig- nificant differences from control; from SQ 22,536 (10-4 M) treat- ment; and from SQ 22,536 (10-4 M) plus beraprost (10-5 M) treat- ment (one-way ANOVA). C Comparison between elevation of cyclic AMP content and relaxation in guinea-pig aorta. Basal lev- els of cyclic AMP content (2.5±0.5 pmol per mg protein, n=6 in duplicates) were not subtracted from the total. Percent relaxation values are the total mean values of all preparations used in the pre- sent study. In the case of beraprost, less cyclic AMP was needed to produce equivalent relaxation as observed with forskolin. Bera- prost (10-5 M; n=6 for cyclic AMP and n=43 for tension); Bera- prost (10-5 M) + SQ 22,536 (10-4 M; n=5 for cyclic AMP and n=10 Fig. 7 Iberiotoxin is able to inhibit the beraprost-induced relax-
for tension); Forskolin (10-5 M) (n=2 for cyclic AMP and n=18 for
ation of guinea-pig aorta in the presence of SQ 22,536. Vascular
relaxation is expressed as percent inhibition against the tension de-velopment due to PGF2α (10-5 M) just before administration of be-
self did not significantly affect the basal level of cyclic
raprost. Mean absolute values of the tension development just be-fore addition of beraprost are 2.10±0.15 g (n=7; ❍, SQ 22,536)
AMP content. A comparison of beraprost-induced relax-
and 2.43±0.23 g (n=8; ●, SQ 22,536 plus Iberiotoxin). Data are
ation with the levels of cyclic AMP (in the absence and
means ± SEM. *P<0.05, **P<0.01 significant differences between
presence of SQ 22,536) clearly shows that beraprost is
two groups (SQ 22,536 10-4 M, Iberiotoxin 10-7 M)
by CTX was also observed in the presence of the adenylylcyclase inhibitor SQ 22,536 (10-4 M), supporting the viewthat Gs can cause relaxation independent of cyclic AMP. As control experiment, CTX was boiled for 20 min at100°C, which practically eliminated its relaxant effect. Todetermine the role of MaxiK channels in this CTX-in-duced relaxation, Iberiotoxin was added after CTX treat-ment. CTX-induced relaxation was significantly counter-acted by Iberiotoxin (10-7 M) in the presence or absenceof SQ 22,536 (10-4 M; Fig. 8B). To rule out the possibilitythat tension decrement in the presence of CTX is only dueto spontaneous muscle fatigue and not due to Gs-mediatedrelaxation, we performed experiments preincubating themuscle with CTX prior PGF2α and Iberiotoxin treatments. Similar results as those shown in Fig. 8A,B were obtained(n=4 to eight experiments). In conclusion, activation of theCTX-sensitive G protein, Gs, is able to induce a signifi-cant (~60%) muscle relaxation of guinea-pig aorta thatcan be explained in part by activation of MaxiK channelindependent of cyclic AMP-mediated mechanisms. Fig. 8 A, B Cholera toxin (CTX), a stimulator of Gs, produces an Iberiotoxin-sensitive relaxation of guinea-pig aorta in the presence or absence of the adenylyl cyclase inhibitor SQ 22,536. A Relax-
ation by CTX (1 µg/ml, 6-h incubation) in the muscle contractedwith PGF2α (10-5 M). Vascular relaxation is expressed as percent
The present findings indicate that activation of MaxiK
2α-induced contraction just before CTX ad-
ministration. CTX induced similar vascular relaxation in the pres-
channels plays a major role in mediating the relaxation in-
ence or absence of SQ 22,536 (SQ, 10-4 M). Boiling (100°C,
duced by stimulation of smooth muscle IP receptors in
20 min) CTX prevented its relaxant effect. Mean absolute values
guinea-pig aorta. We also provide evidence for a dual
of the tension development by PGF2α (10-5 M) after 1-h incubation
mechanism of action for IP receptor stimulation by be-
are: 2.40±0.36 g (n=4; control), 2.53±0.30 g (n=3; SQ 22,536 10-4 M), 2.33±0.20 g (n=10; CTX 1 µg/ml), 2.43±0.34 g (n=5; SQ
raprost including both cyclic AMP-dependent and -inde-
22,536 10-4 M plus CTX 1 µg/ml), and 2.11±0.11 g (n=5; CTX,
pendent (via Gs) pathways, which facilitate MaxiK chan-
boiled). Note that SQ 22,536 was applied prior (20 min) contrac-
nel activity leading to vascular relaxation.
tion with PGF2α. Data are means ± SEM. **P<0.01 significant dif-
The relaxation of guinea-pig aorta in response to be-
ferences from control group. B Iberiotoxin inhibits vascular relax- ation by CTX or CTX plus SQ 22,536. Iberiotoxin (10-7 M) was
raprost is mediated via an activation of plasma-membrane
applied 6 h after the incubation with CTX (1 µg/ml). Data are
K+ channels since the relaxant response to beraprost was
means ± SEM of 5-8 experiments. *P<0.05 significant difference
almost abolished in the presence of high-KCl (Fig. 1). The
driving force for K+ efflux and the subsequent membranehyperpolarization would be considerably reduced by diminu-tion of the chemical gradient in the medium containing
way was inhibited (Fig. 7). These findings strongly indi-
high K+ (Wise and Jones 1996). The main K+ channel in-
cate that the activation of MaxiK channel contributes at
volved in beraprost-induced relaxation is the MaxiK
least by ~50% to the cyclic AMP-independent component
channel because of the following observations: (1) Iberi-
otoxin (10-7 M) dramatically diminished beraprost-in-duced relaxation close to the level obtained in high-KClsolution (Fig. 2 vs. Fig. 1). MaxiK channel is not inhibited
Activation of Gs with cholera toxin relaxes guinea-pig
by glibenclamide (10-5 M), apamin (10-7 M), 4-aminopy-
aorta in a cyclic AMP-independent manner
ridine (10-2 M; Wallner et al. 1995), but is blocked selec-
tively by Iberiotoxin (Galvez et al. 1990); (2) Beraprost-induced relaxation was also inhibited by TEA at a con-
To test the possibility that beraprost-induced cyclic AMP-
centration that is considered to block MaxiK channels
independent activation of MaxiK channels occurs via a di-
(<2×10-3 M; Nelson and Quayle 1995; Wallner et al. 1995),
rect G protein channel stimulation, we first investigated
and the inhibition was similar to that observed in the pres-
whether cholera toxin (CTX), a stimulator of Gs, could
ence of Iberiotoxin (Fig. 2); (3) The relaxation induced by
mimic the relaxant effect of beraprost. CTX (1 µg/ml)
beraprost was not affected by glibenclamide (10-6 M),
was applied to the bath solution after the contraction due
which as expected almost abolished the relaxation due to
to PGF2α (10-5 M) reached a steady-state level, and was
cromakalim (Fig. 3). Thus, although the KATP channel has
maintained in the bath for 6 h in the continued presence of
a significant contribution to the vascular relaxation in-
PGF2α. As shown in Fig. 8A, CTX by itself significantly
duced by PGI2 analogues in some blood vessels (Jackson
relaxed PGF2α-contracted vessels. Moreover, relaxation
et al. 1993; Lombard et al. 1999; Schubert et al. 1997), in
guinea-pig aorta the participation of this class of K+ chan-
dent mechanism(s) to the overall muscle relaxation is def-
nel can be ruled out; and (4) low concentrations of Ba2+
initely important. A significant role of cyclic AMP-inde-
(10-5 M) did not affect beraprost-induced relaxation pendent pathway(s) in the regulation of muscle tone was(Fig. 3), indicating that an inward rectifier type of K+
also implied in the vascular relaxations induced by other
channel (Bradley et al. 1999) may also be ruled out as the
PGI2 analogues, iloprost and cicaprost, and by a β-adreno-
main K+ channel involved in the relaxation induced by be-
ceptor agonist, isoprenaline (Clapp et al. 1998; Kume et
al. 1994; Scornik et al. 1993; Turcato and Clapp 1999).
Both Iberiotoxin (10-7 M) and TEA (2×10-3 M) did not
A cyclic AMP-independent mechanism responsible for
completely eliminate the overall relaxant effect of be-
the activation of MaxiK channel has been shown with
raprost, whereas the relaxation was almost abolished in
β-adrenoceptor stimulation in the smooth muscle from
high-KCl solution. This implies that a small portion of be-
coronary artery (Scornik et al. 1993) and airway muscle
raprost-induced relaxation can be ascribed to a K+ channel
(Kume et al. 1994). In these reports, direct stimulatory ac-
other than the MaxiK channel. This non-MaxiK channel-
tion of the guanine nucleotide regulatory protein, Gs, un-
mediated relaxant component may be attributed to Kv
derlies the cyclic AMP-independent modulation of the
channel since 4-AP (10-4 M) inhibited beraprost-induced
MaxiK channel (Kume et al. 1994; Scornik et al. 1993).
relaxation to a small degree. The extent of the inhibition
The cell surface IP receptor is known to be coupled to
by 4-AP (~15%; Fig. 3) was close to the relaxant compo-
adenylyl cyclase via Gs (Leffler 1997; Narumiya et al.
nent remaining in the presence of Iberiotoxin (~19%) or
1999; Wise and Jones 1996). Thus, IP receptor stimula-
TEA (~25%) at 3×10-5 M beraprost. Indeed, the modula-
tion may activate the MaxiK channel through a direct Gs-
tion of Kv channel(s) (4-aminopyridine-sensitive, delayed
protein coupling in guinea-pig aortic smooth muscle cells.
rectifier K+ channels) by cyclic AMP elevating stimuli in
Consistent with this idea, in the present study we showed
vascular smooth muscle cells has been suggested (Cole et
that CTX (1 µg/ml), an activator of the stimulatory G pro-
al. 1996; Standen and Quayle 1998). Nevertheless, the
tein of adenylyl cyclase (Gs), substantially relaxed PGF2α-
main K+ channel contributing to the overall relaxation due
induced muscle contraction both in the absence and pres-
to beraprost seems to be the MaxiK channel in guinea-pig
ence of SQ 22,536 (10-4 M; Fig. 8A). Moreover, the re-
aorta. Since we used endothelium-denuded preparations
laxation due to CTX was partially but significantly coun-
in this study, a possible participation of endothelial K+
teracted by Iberiotoxin (10-7 M; Fig. 8B). These mechani-
cal studies strongly suggest that activation of Gs-protein
As mentioned above, another important implication of
could produce relaxation of guinea-pig aorta through di-
the present findings is that cyclic AMP-independent mech-
rect modulation of smooth muscle cell MaxiK channels.
anism(s) significantly (but not exclusively) contributes to
In addition, several lines of evidence indicate that IP re-
MaxiK channel-mediated relaxation induced by beraprost
ceptors can be coupled to non-Gs type of G-proteins in-
in guinea-pig aorta. This conclusion is based on the fol-
cluding Gi, Go and Gp (Leffler 1997; Narumiya et al. 1999;
lowing observations: (1) Similar to the iloprost-induced
Wise and Jones 1996); thus, possible coupling with these
relaxation (Turcato and Clapp 1999), the relaxation in-
additional pathways needs to be investigated.
duced by beraprost was not affected by the adenylyl cy-
Our present data clearly show that IP receptor activa-
clase inhibitor, SQ 22,536 (10-4 M); although the same
tion by beraprost elevates cyclic AMP contents in guinea-
concentration of SQ 22,536 prevented the increase in
pig aortic smooth muscle cells (Fig. 6), and that additional
cyclic AMP content by this IP agonist. This pharmaco-
cyclic AMP-dependent mechanisms are partially involved
mechanical and biochemical evidence indicates that be-
in beraprost induced-relaxation. Elevated cyclic AMP
raprost is able to relax guinea-pig aorta even when the
can produce MaxiK channel-mediated muscle relaxation
adenylyl cyclase is blocked, and thus, implies the exis-
(Fig. 5), and RO-20-1724 (cyclic AMP-specific phospho-
tence of cyclic AMP-independent mechanism(s) responsi-
diesterase inhibitor) can potentiate the beraprost-induced
ble for MaxiK channel-mediated vascular relaxation; (2)
relaxation in an Iberiotoxin-sensitive manner (Fig. 4). In
Iberiotoxin blocked about 50% of beraprost-induced re-
agreement with this view, 8-bromo-cyclic AMP-induced
laxation in the presence of SQ 22,536; whereas it blocked
relaxation was significantly suppressed by Iberiotoxin ap-
~80% in the absence of this adenylyl cyclase inhibitor;
plied either before or after stimulation with this cyclic
and (3) the level of cyclic AMP needed to elicit the same
AMP analogue. Furthermore, a complete counteracting
amount of vascular relaxation was much less for beraprost
action of Iberiotoxin was obtained for forskolin-induced
than for forskolin. This was more evident when the
relaxation when this channel blocker was applied during
adenylyl cyclase was inhibited with SQ 22,536 (Fig. 6C).
the muscle relaxation. However, and as previously re-
Although the compartmentalization of cyclic AMP neces-
ported (Clapp et al. 1998), if Iberiotoxin was pre-applied
sary for the regulation of muscle tone has been postulated
to block forskolin-induced relaxation of guinea-pig aorta,
(Laurenza et al. 1989), our finding may indicate that ele-
the channel blocker was unable to prevent this relaxation;
vation of cyclic AMP itself cannot efficiently generate
at present, we do not have an explanation for this phe-
vascular smooth muscle relaxation. Since the concentra-
tion-response relationships for the relaxant effect of be-
In contrast to guinea-pig aorta, in rat tail artery smooth
raprost were almost identical in the absence and presence
muscle a cyclic AMP-dependent mechanism has a sub-
of SQ 22,536, the contribution of cyclic AMP-indepen-
stantial contribution in the MaxiK channel-mediated mus-
cle relaxation after IP receptor stimulation (Schubert et al. Acknowledgements This study was supported in part by Grant-
1996, 1997). Mechanisms proposed for the explanation of
in-Aid (12672226) from the Ministry of Education, Science,Sports, and Culture, Japan (Y.T.), the Nestle Foundation (Y.T.),
cyclic AMP-dependent MaxiK channel activation are: (1)
the Pharmacological Research Foundation, Tokyo (Y.T.), and NIH
phosphorylation by cyclic AMP-dependent protein kinase
(PKA; Kume et al. 1989; Meera et al. 1995; Schubert etal. 1996; Scornik et al. 1993); (2) PKA-triggered increasein Ca2+ spark frequency through ryanodine-sensitive Ca2+
release channels (Porter et al. 1998); and (3) modulation
Adeagbo ASO, Malik KU (1990) Mechanism of vascular actions
by cyclic GMP-dependent protein kinase via cyclic AMP
of prostacyclin in the rat isolated perfused mesenteric arteries.
(Lincoln and Cornwell 1993). However, experiments us-
ing the soluble guanylyl cyclase inhibitor, ODQ (10-5 M),
Akiba T, Miyazaki M, Toda N (1986) Vasodilator actions of TRK-
showed that beraprost-induced relaxation was not affected
100, a new prostaglandin I2 analogue. Br J Pharmacol 89:703-711
by this compound (not shown). Thus, a possible participa-
Bradley KK, Jaggar JH, Bonev AD, Heppner TJ, Flynn ER, Nel-
tion of cyclic GMP as a mechanism involved in the relax-
son MT, Horowitz B (1999) Kir2.1 encodes the inward rectifier
ation due to beraprost seems unlikely.
potassium channel in rat arterial smooth muscle cells. J Physiol
Beraprost exhibited an additional contraction at con-
centration ranges over 10-6 M in the muscle contracted with
Clapp LH, Turcato S, Hall S, Baloch M (1998) Evidence that Ca2+-
activated K+ channels play a major role in mediating the vascu-
high-KCl. Thus, high concentrations of IP agonists may
lar effects of iloprost and cicaprost. Eur J Pharmacol 356:
produce vascular contraction as well as relaxation in
guinea-pig aorta. The contractile effect of beraprost is pre-
Cocks TM, King SJ, Angus JA (1990) Glibenclamide is a compet-
dominant in 80 mM KCl-contracted muscle since the
itive antagonist of the thromboxane A2 receptor in dog coro-nary artery in vitro. Br J Pharmacol 100:375-378
counteracting relaxant component associated with K+
Cole WC, Clement-Chomienne O, Aiello EA (1996) Regulation of
channel activity is minimized in high-KCl-containing me-
4-aminopyridine-sensitive, delayed rectifier K+ channels in
dium. Although the causal relationship between the ten-
vascular smooth muscle by phosphorylation. Biochem Cell
sion development and elevation of cyclic AMP following
Delaey C, Van de Voorde J (1995) Prostanoid-induced contrac-
stimulation with beraprost is not clear at present, genera-
tions are blocked by sulfonylureas. Eur J Pharmacol 280:179-
tion of muscle tone by PGI2 with concomitant cyclic AMP
elevation has also been reported in rabbit aorta (Vegesna
Dumas M, Dumas JP, Rochette L, Advenier C, Giudicelli JF
and Diamond 1986). Concerning the vascular contraction
(1997) Role of potassium channels and nitric oxide in the ef-
induced by other IP agonists, iloprost has been implicated
fects of iloprost and prostaglandin E1 on hypoxic vasoconstric-tion in the isolated perfused lung of the rat. Br J Pharmacol
in the production of constrictor prostaglandin(s) and/or
thromboxane A2 (TXA2; Clapp et al. 1998; Ozaki et al.
Galvez A, Gimenez-Gallego G, Reuben JP, Roy-Contancin L,
Feigenbaum P, Kaczorowski J, Garcia ML (1990) Purification
1998), which would counterbalance its relaxant effect in
and characterization of a unique, potent, peptidyl probe for thehigh conductance calcium-activated potassium channel from
guinea-pig aorta. However, in our experiments involve-
venom of the scorpion Buthus tamulus. J Biol Chem 265:
ment of constrictor prostaglandin(s) (e.g. prostaglandin
Jackson WF, König A, Dambacher T, Busse R (1993) Prostacy-
2, prostaglandin F2α) and/or TXA2 seems unlikely since
beraprost-induced relaxation was not affected by gliben-
clin-induced vasodilation in rabbit heart is mediated by ATP-sensitive potassium channels. Am J Physiol 264:H238-H243
clamide (Fig. 3A), which in other vascular beds inhibits vas-
Katano Y, Endoh M (1990) Differential effects of Ro 20-1724 and
cular contractions induced by these vasoactive substances
isobutylmethylxanthine on the basal force of contraction and
(Cocks et al. 1990; Delaey and Van de Voorde 1995).
beta-adrenoceptor-mediated response in the rat ventricular myo-
In summary, our present findings indicate that endo-
cardium. Biochem Biophys Res Commun 167:123-129
thelium-independent relaxation of guinea-pig aorta in re-
Kukovetz WR, Holzmann S, Wurm A, Poch G (1979) Prostacyclin
increases cAMP in coronary arteries. J Cyclic Nucleotide Res
sponse to the IP agonist, beraprost, is almost totally as-
cribed to the activation of plasma-membrane K+ channels.
Kume H, Takai A, Tokuno H, Tomita T (1989) Regulation of
MaxiK channel is the predominant mediator of vascular
Ca2+-dependent K+-channel activity in tracheal myocytes by
relaxation following stimulation of IP receptor; while K
Kume H, Hall IP, Washabau RJ, Takagi K, Kotlikoff MI (1994)
channel(s) may be modestly involved in the relaxant
Beta-adrenergic agonists regulate KCa channels in airway
mechanism. Contribution of KATP channel can be ruled out
smooth muscle by cAMP-dependent and -independent mecha-
as the mechanism by which beraprost relaxes guinea-pig
aorta. Although cyclic AMP content is significantly ele-
Laurenza A, Sutkowski EM, Seamon KB (1989) Forskolin: a spe-
cific stimulator of adenylate cyclase or a diterpene with multi-
vated by beraprost in this vascular tissue, the opening of
ple sites of action? Trends Pharmacol Sci 10:442-447
MaxiK channel dependent on the elevation of intracellular
Leffler CH (1997) Prostanoids: intrinsic modulators of cerebral
cyclic AMP is not overwhelming; whereas the cyclic
AMP-independent component seems to be the major con-
Lincoln TM, Cornwell TL (1993) Intracellular cyclic GMP recep-
tributor of the total relaxation induced by this IP agonist.
Lombard JH, Liu Y, Fredricks KT, Bizub DM, Roman RJ, Rusch
The mechanism of cyclic AMP-independent relaxation is
NJ (1999) Electrical and mechanical responses of rat middle
due in part to the direct activation of MaxiK channels by
cerebral arteries to reduced PO2 and prostacyclin. Am J Phys-
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein
Schubert R, Serebryakov VN, Mewes H, Hopp H-H (1997) Ilo-
measurement with the folin phenol reagent. J Biol Chem 193:
prost dilates rat small arteries: role of KATP- and KCa-channel
activation by cAMP-dependent protein kinase. Am J Physiol
Meera P, Anwer K, Monga M, Oberti C, Stefani E, Toro L, San-
born BM (1995) Relaxin stimulates myometrial calcium-acti-
Scornik FS, Codina J, Birnbaumer L, Toro L (1993) Modulation of
vated potassium channel activity via protein kinase A. Am J
coronary smooth muscle KCa channels by Gsα independent of
phosphorylation by protein kinase A. Am J Physiol 265:H1460-
Miller OV, Aiken JW, Hemker DP, Shebuski RJ, Gorman RR
(1979) Prostacyclin stimulation of dog arterial cyclic AMP lev-
Siegel G, Carl A, Adler A, Stock G (1989) Effect of the prostacy-
clin analogue iloprost on K+ permeability in the smooth muscle
Moncada S, Herman AG, Higgs EA, Vane JR (1977) Differential
cells of the canine carotid artery. Eicosanoids 2:213-222
formation of prostacyclin (PGX or PGI2) by layer of the arter-
Standen NB, Quayle JM (1998) K+ channel modulation in arterial
ial wall. An explanation for the anti-thrombotic properties of
smooth muscle. Acta Physiol Scand 164:549-557
vascular endothelium. Thromb Res 11:323-344
Tanaka Y, Igarashi T, Takayanagi K, Yamaki F, Imai T, Tanaka H,
Murata T, Murai T, Kanai T, Ogaki Y, Sanai K, Kanda H, Sato S,
Shigenobu K (2000) Comparison of pharmacological charac-
Kajikawa N, Umetsu T, Matsuura H, Fukatsu Y, Isogaya M,
teristics of noradrenaline- and thapsigargin-activated Ca2+ en-
Yamada N, Nishio S (1989) General pharmacology of be-
try channels responsible for mechanical responses of guinea-
raprost sodium. 2nd communication: effect on the autonomic,
pig aorta. Naunyn-Schmiedeberg's Arch Pharmacol 362:160-
cardiovascular and gastrointestinal systems, and other effects.
Toro L, Wallner M, Meera P, Tanaka Y (1998) Maxi-KCa, a unique
Narumiya S, Sugimoto Y, Ushikubi F (1999) Prostanoid receptors:
member of the voltage-gated K channel superfamily. News
structures, properties, and functions. Physiol Rev 79:1193-
Turcato S, Clapp LH (1999) Effects of the adenylyl cyclase in-
Nelson MT, Quayle JM (1995) Physiological roles and properties
hibitor SQ22536 on iloprost-induced vasorelaxation and cyclic
of potassium channels in arterial smooth muscle. Am J Physiol
AMP elevation in isolated guinea-pig aorta. Br J Pharmacol
Ozaki H, Abe A, Uehigashi Y, Kinoshita M, Hori M, Mitsui-Saito
Vegesna RV, Diamond J (1986) Elevation of cyclic AMP by
M, Karaki H (1996) Effects of a prostaglandin I2 analog ilo-
prostacyclin is accompanied by relaxation of bovine coronary
prost on cytoplasmic Ca2+ levels and muscle contraction in iso-
arteries and contraction of rabbit aortic rings. Eur J Pharmacol
lated guinea pig aorta. Jpn J Pharmacol 71:231-237
Porter VA, Bonev AD, Knot HJ, Heppner TJ, Stevenson AS, Klep-
Wallner M, Meera P, Ottolia M, Kaczorowski GJ, Latorre R, Gar-
pisch T, Lederer WJ, Nelson MT (1998) Frequency modulation
cia ML, Stefani E, Toro L (1995) Characterization of and mod-
of Ca2+ sparks is involved in regulation of arterial diameter by
ulation by a β-subunit of a human maxi KCa channel cloned
cyclic nucleotides. Am J Physiol 274:C1346-C1355
from myometrium. Receptors Channels 3:185-199
Schubert R, Serebryakov VN, Engel H, Hopp H-H (1996) Iloprost
Wise H, Jones RL (1996) Focus on prostacyclin and its novel
activates KCa channels of vascular smooth muscle cells: role of
mimetics. Trends Pharmacol Sci 17:17-21
cAMP-dependent protein kinase. Am J Physiol 271:C1203-C1211
International IBD Genetics Consortium Thiopurine Induced Leucopaenia Case Report Form On completion, please return to: Thiopurine Induced Leucopaenia Introduction Please complete all boxes where indicated and in black ball point pen. If you make a mistake please put a line through the box, initial and date and write answer to the side. The patient identification number is th
Olaf-Georg Issinger: List of publications 138. Persson, T., Yde, C.W., Rasmussen, J.E., Rasmussen, T.L., Guerra, B., Issinger, O.-G., and NielsenJ. (2007) Carboxyl and arboxamide pyrazoles as protein kinase inhibitors in corrupt eukaryotic signaltransduction: Induction of growth arrest in MCF-7 cancer cel s. Organic & Bioorganic Chemistry, in press137. Yde, C.W., Frogne, T, Lykkesfeldt,
 At concentration ranges over 10-6 M, a small contractioninstead of relaxation was attained by beraprost (Fig. 1B).
At concentration ranges over 10-6 M, a small contractioninstead of relaxation was attained by beraprost (Fig. 1B).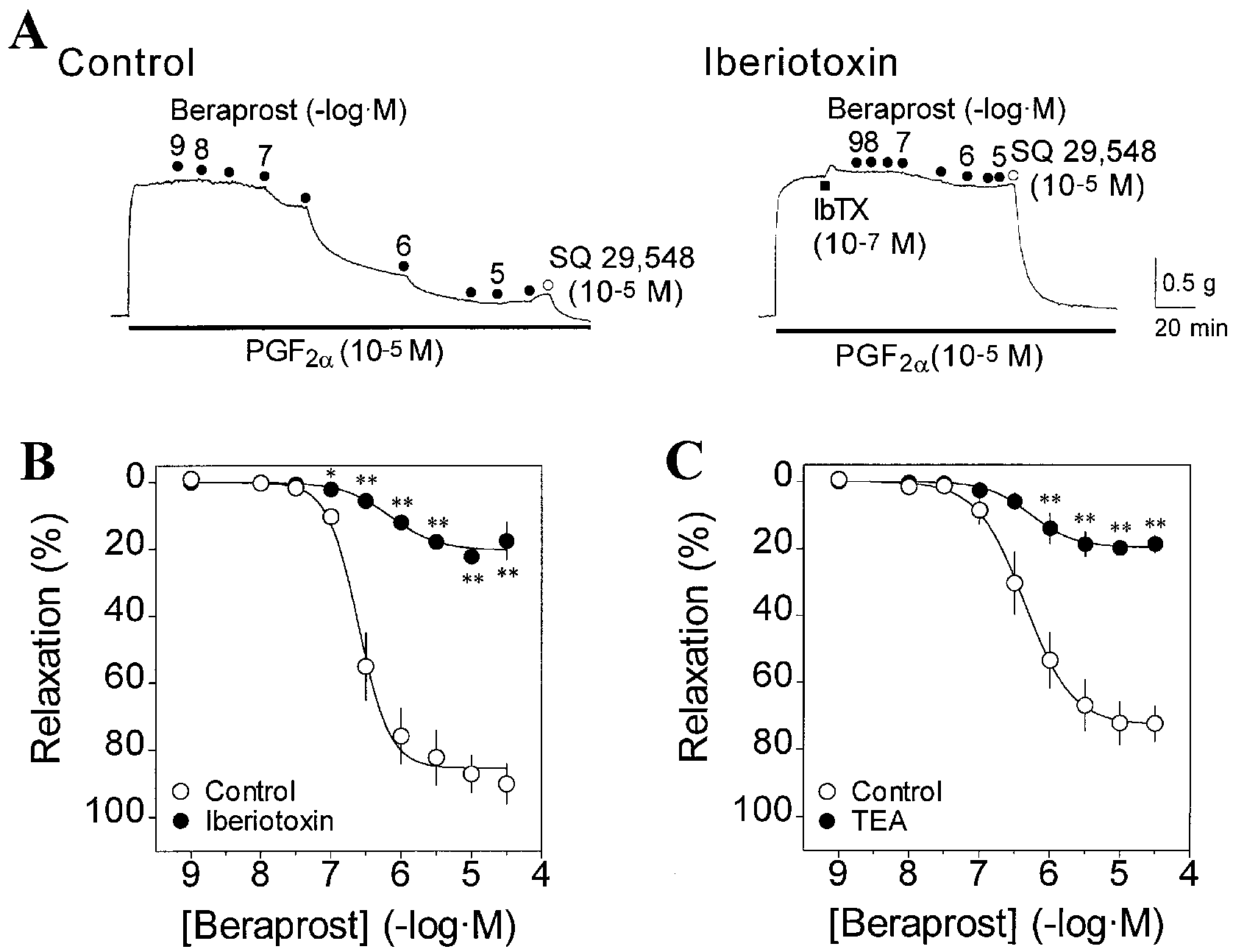 Table 1 Effects of high-KCl and MaxiK channel blockers on the
Table 1 Effects of high-KCl and MaxiK channel blockers on the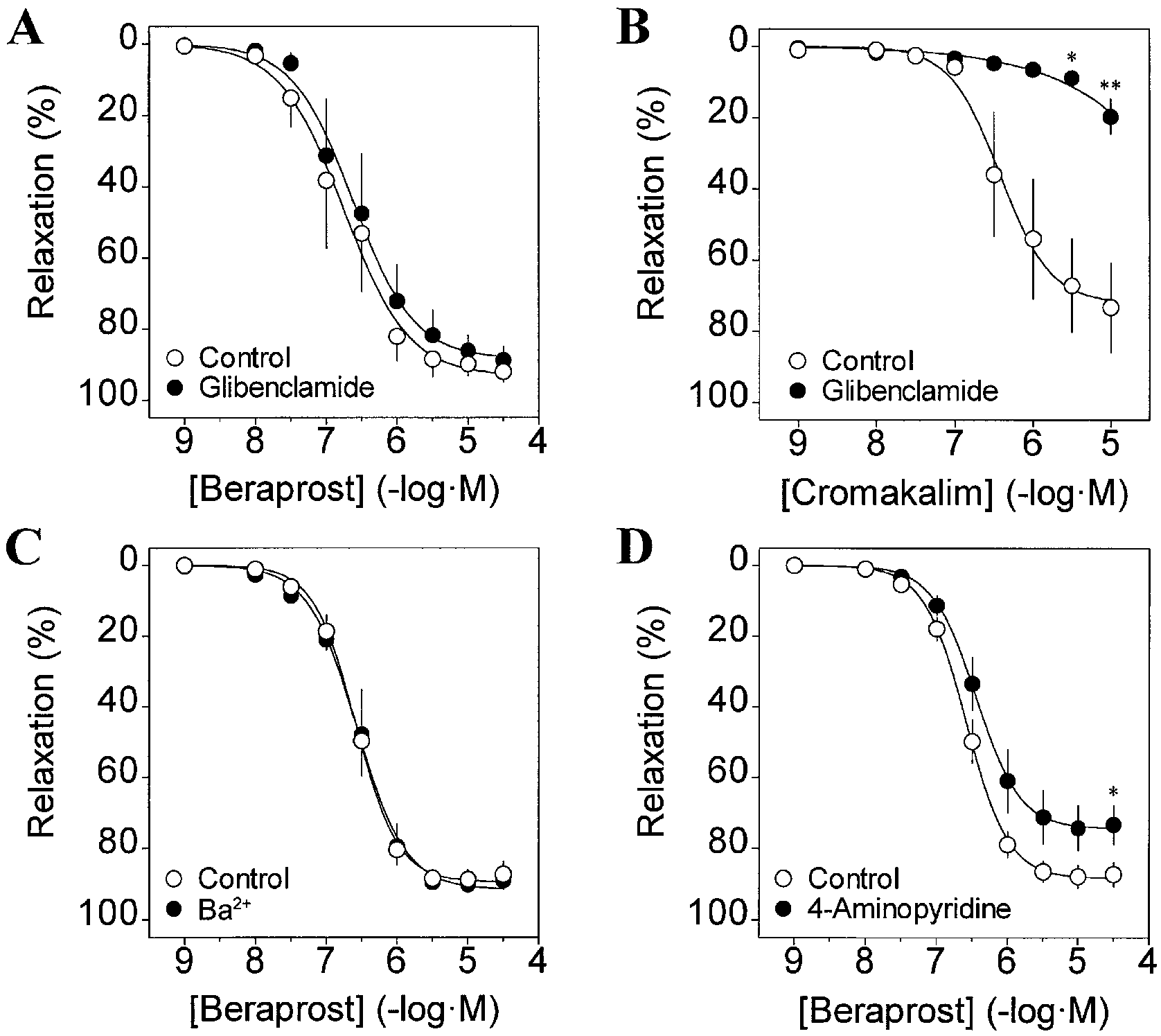 Fig. 3A-D 4-Aminopyridine (4-AP) but not glibenclamide or
Fig. 3A-D 4-Aminopyridine (4-AP) but not glibenclamide or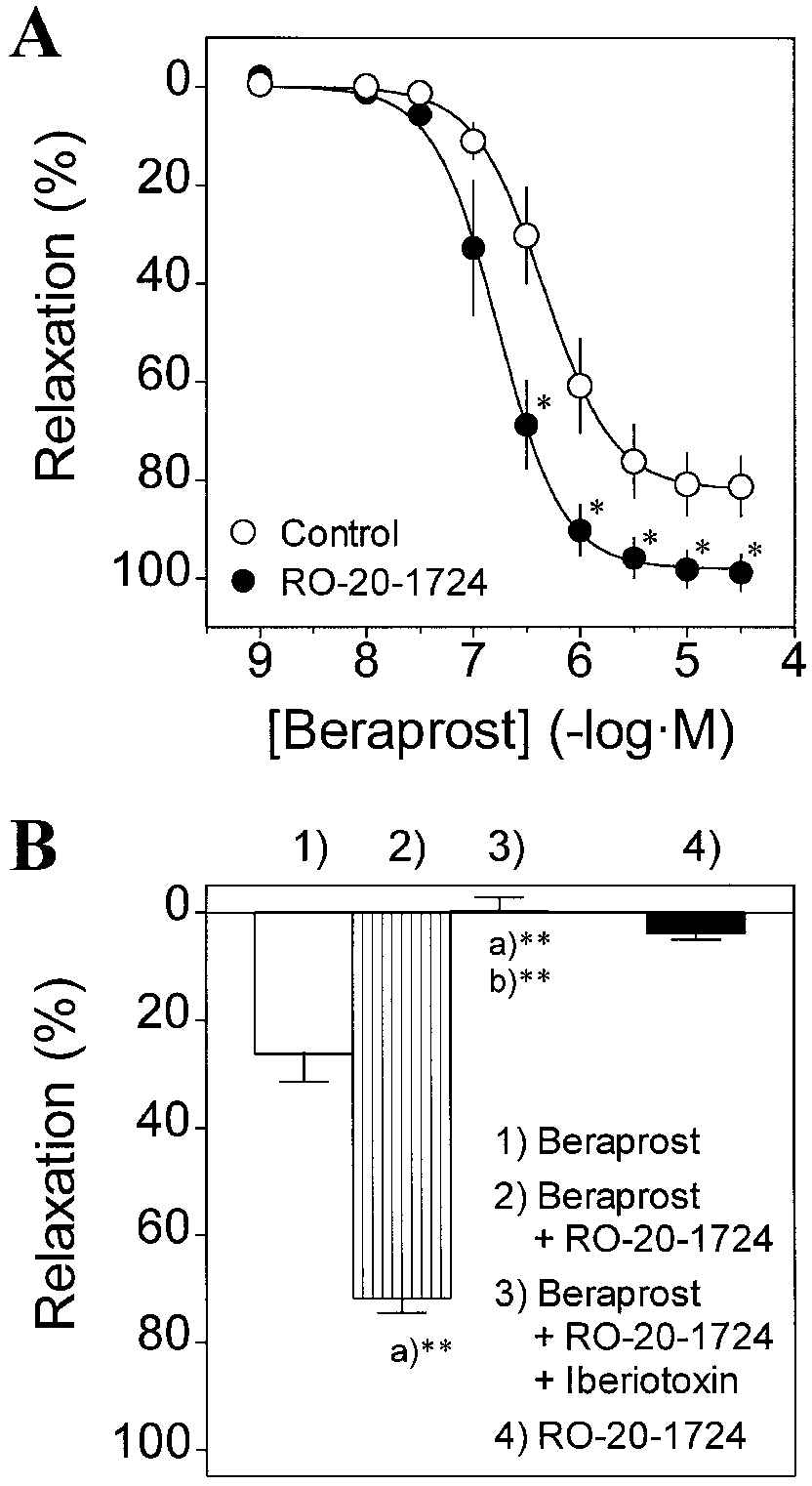 Fig. 4B). Note that muscle tension decrement due to RO-20-1724 (10-4 M) alone was only 3.9±1.2% (n=3).
Fig. 4B). Note that muscle tension decrement due to RO-20-1724 (10-4 M) alone was only 3.9±1.2% (n=3).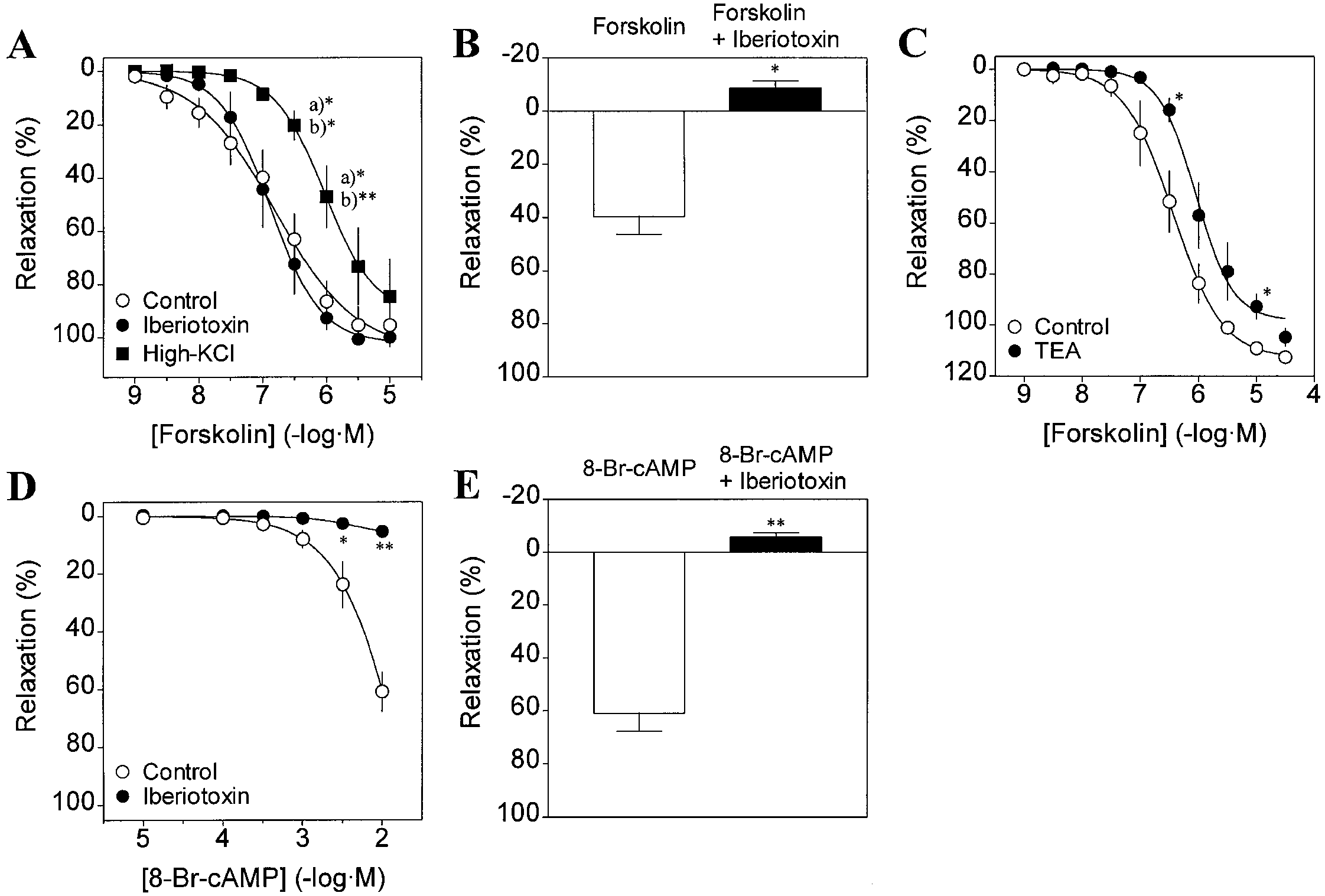 Fig. 5A-E MaxiK channel blockers also inhibit the relaxations in-
Fig. 5A-E MaxiK channel blockers also inhibit the relaxations in-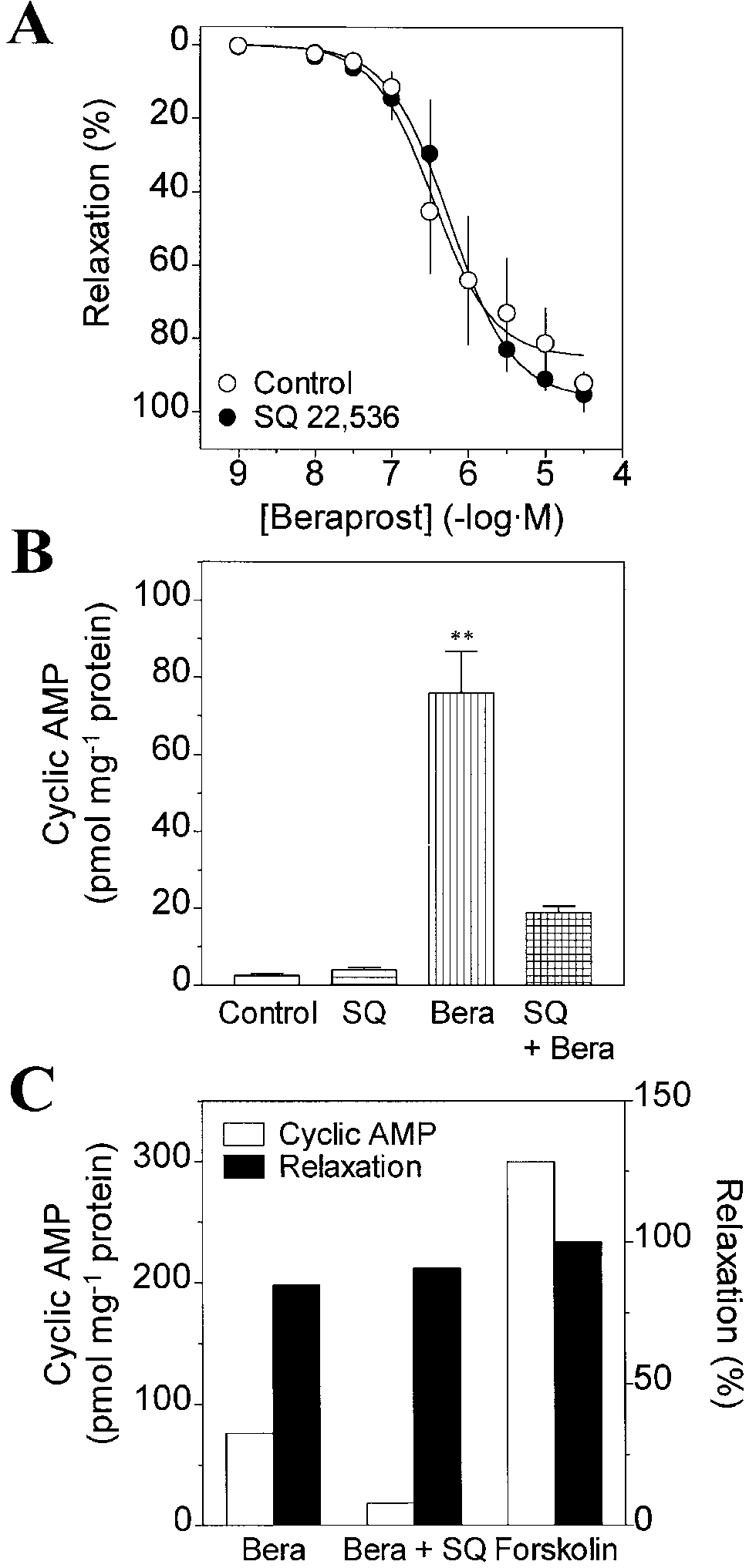
 able to relax the vascular muscle even when the adenylylcyclase activity is strongly inhibited by SQ 22,536 (Fig. 6C).
able to relax the vascular muscle even when the adenylylcyclase activity is strongly inhibited by SQ 22,536 (Fig. 6C).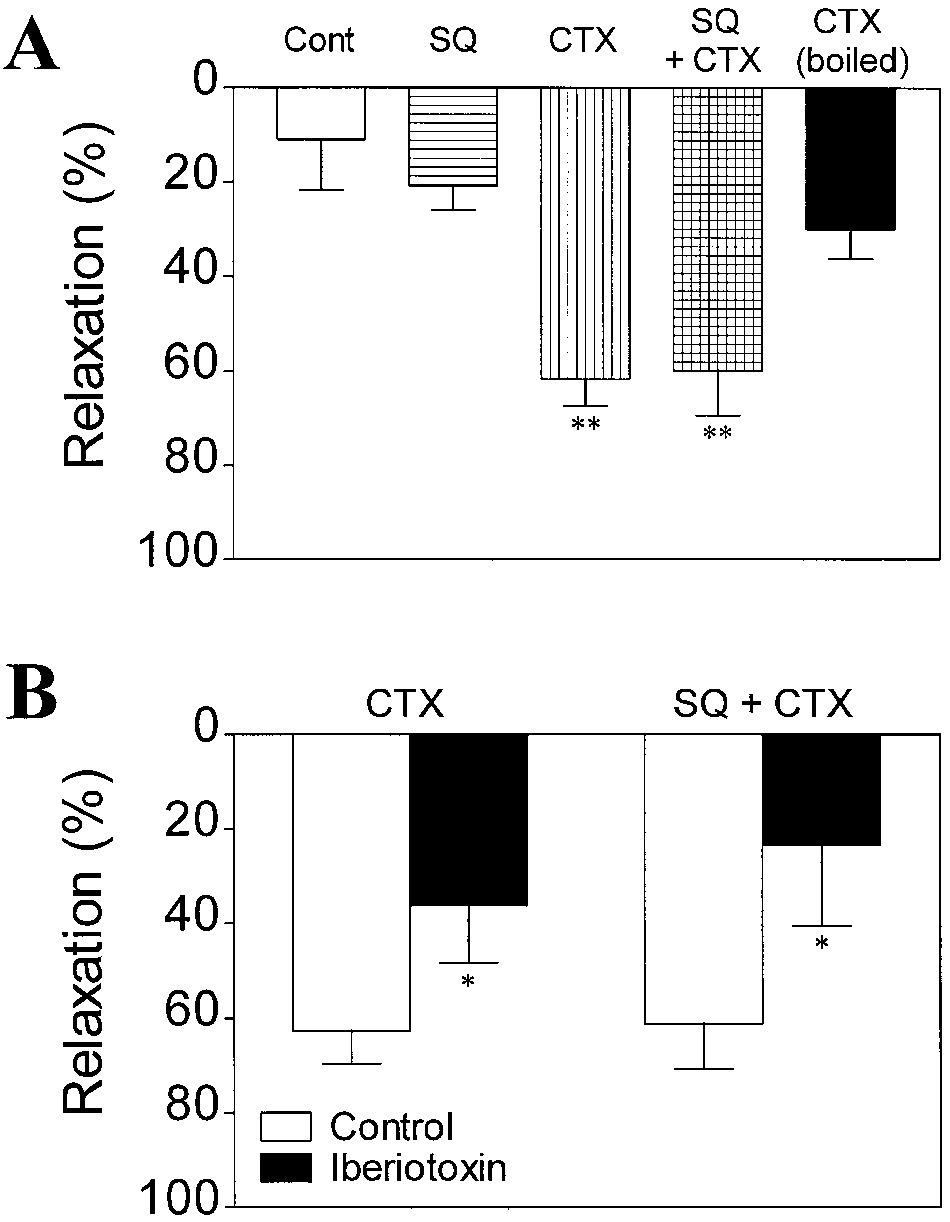 by CTX was also observed in the presence of the adenylylcyclase inhibitor SQ 22,536 (10-4 M), supporting the viewthat Gs can cause relaxation independent of cyclic AMP.
by CTX was also observed in the presence of the adenylylcyclase inhibitor SQ 22,536 (10-4 M), supporting the viewthat Gs can cause relaxation independent of cyclic AMP.