L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
The growth response of plants to elevated co2 under non-optimal conditions
Oecologia (2001) 129:1-20DOI 10.1007/s004420100736
Hendrik Poorter. Marta Pérez-Soba
The growth response of plants to elevated CO under non-optimal
Received: 10 August 2000 / Accepted: 19 April 2001 / Published online: 25 July 2001 Springer-Verlag 2001
Abstract Under benign environmental conditions, plant
and decrease in others, resulting in an average interac-
growth is generally stimulated by elevated atmospheric
tion with elevated CO that was not significant. Under
CO concentrations. When environmental conditions be-
high ozone concentrations, the relative growth enhance-
come sub- or supra-optimal for growth, changes in the
ment by elevated CO was strongly increased, to the ex-
biomass enhancement ratio (BER; total plant biomass at
tent that high CO even compensated in an absolute way
elevated CO divided by plant biomass at the current
for the harmful effect of ozone on growth. No systematic
CO level) may occur. We analysed literature sources
difference in response was found between herbaceous
that studied CO ×environment interactions on the growth
and woody species for any of the environmental vari-
of herbaceous species and tree seedlings during the vege-
tative phase. For each experiment we calculated the dif-ference in BER for plants that were grown under 'opti-
Keywords Nutrients. Water. Light. Temperature. Salt.
mal' and 'non-optimal' conditions. Assuming that inter-
actions would be most apparent if the environmentalstress strongly diminished growth, we scaled the differ-ence in the BER values by the growth reduction due to
The complex effect of elevated CO on plant growth
the stress factor. In our compilation we found a largevariability in CO ×environment interactions between ex-
The current increase in the atmospheric CO concentra-
periments. To test the impact of experimental design, we
tion has triggered a wide variety of botanical investiga-
simulated a range of analyses with a plant-to-plant varia-
tions during the last two decades, at a range of integra-
tion in size common in experimental plant populations,
tion levels. Notwithstanding this huge effort, we still
in combination with a number of replicates generally
have only a limited understanding about the effect of ele-
used in CO ×environment studies. A similar variation in
vated CO on plant growth. There is considerable varia-
results was found as in the compilation of real experi-
tion in the direction and magnitude of growth responses
ments, showing the strong impact of stochasticity. We
to elevated CO , partly depending on the duration of the
therefore caution against strong inferences derived from
exposure, plant development, species (e.g. species that
single experiments and suggest rather a reliance on aver-
differ in inherent growth rate or type of photosynthetic
age interactions across a range of experiments. Averaged
pathway) and the availability of primary resources
over the literature data available, low soil nutrient supply
(Kimball 1986a; Idso and Idso 1994; Poorter et al. 1996;
or sub-optimal temperatures were found to reduce the
Curtis and Wang 1998; Saxe et al. 1998). However, there
proportional growth stimulation of elevated CO. In con-
is still debate about when and where and to what extent
trast, BER increased when plants were grown at low wa-
these factors are important (Kimball 1986b; Idso and
ter supply, albeit relatively modestly. Reduced irradiance
Idso 1994; Lloyd and Farquhar 1996, 2000; Poorter
or high salinity caused BER to increase in some cases
1998; Stitt and Krapp 1999). The situation becomes evenmore complex if we take into account that concomitantwith the increased level of CO , there are also increases
in the level of air pollutants (ozone, nitrogen oxides, sul-
Plant Ecophysiology, Utrecht University, P.O. Box 800.84,
phur dioxide) and ultraviolet radiation. Enhanced deposi-
3508 TB Utrecht, The Netherlandse-mail: h.poorter@bio.uu.nl
tion of air pollutants results in eutrophication and acidifi-
Tel.: +31-30-2536859, Fax: +31-30-2518366
cation of natural ecosystems. Increased emissions ofCO , methane and chlorofluorocarbons might result in
Alterra, P.O. Box 47, 6700 AA Wageningen, The Netherlands
increased temperature and alterations in other climate
parameters, such as the distribution and intensity of
stress was then calculated as the reduction in total biomass at am-
clouds (light) and precipitation (water). We therefore
bient CO of plants grown at the non-optimal level compared to
the total biomass of plants grown at the optimal level. We call this
need to analyse how these changing environmental fac-
the 'growth reduction due to stress' (GRS) and calculated it as:
tors may modify the impact of elevated CO on plant
A range of research papers and reviews has dealt with
the interactions between elevated atmospheric CO con-
with M and M being the total biomass of plants at the optimal
level O and at a certain sub- or supra-optimal level S, respectively.
centration and environmental factors (e.g. Kimball
We thereby assume that the higher the GRS, i.e. the larger the dif-
1986b; Gifford 1992; Idso and Idso 1994; Curtis and
ference in biomass between the optimal and a non-optimal level,
Wang 1998; Poorter 1998; Luo and Mooney 1999). In
the stronger was the stress experienced by the plants. Because ra-
most experiments, the CO effect is analysed at two lev-
tios are ln-normally distributed by nature, we first ln-transformed
the BER values obtained under optimal and non-optimal condi-
els of another environmental factor, sometimes with
tions, and then scaled the difference between these two values by
quite contrasting results that hinder generalisations
the growth reduction observed because of the interacting stress
across experiments (Rawson 1992). Differences in re-
sponse between species might be responsible for differ-ent results. Far less attention has been paid to the possi-
bility that these differences are merely due to chance. In
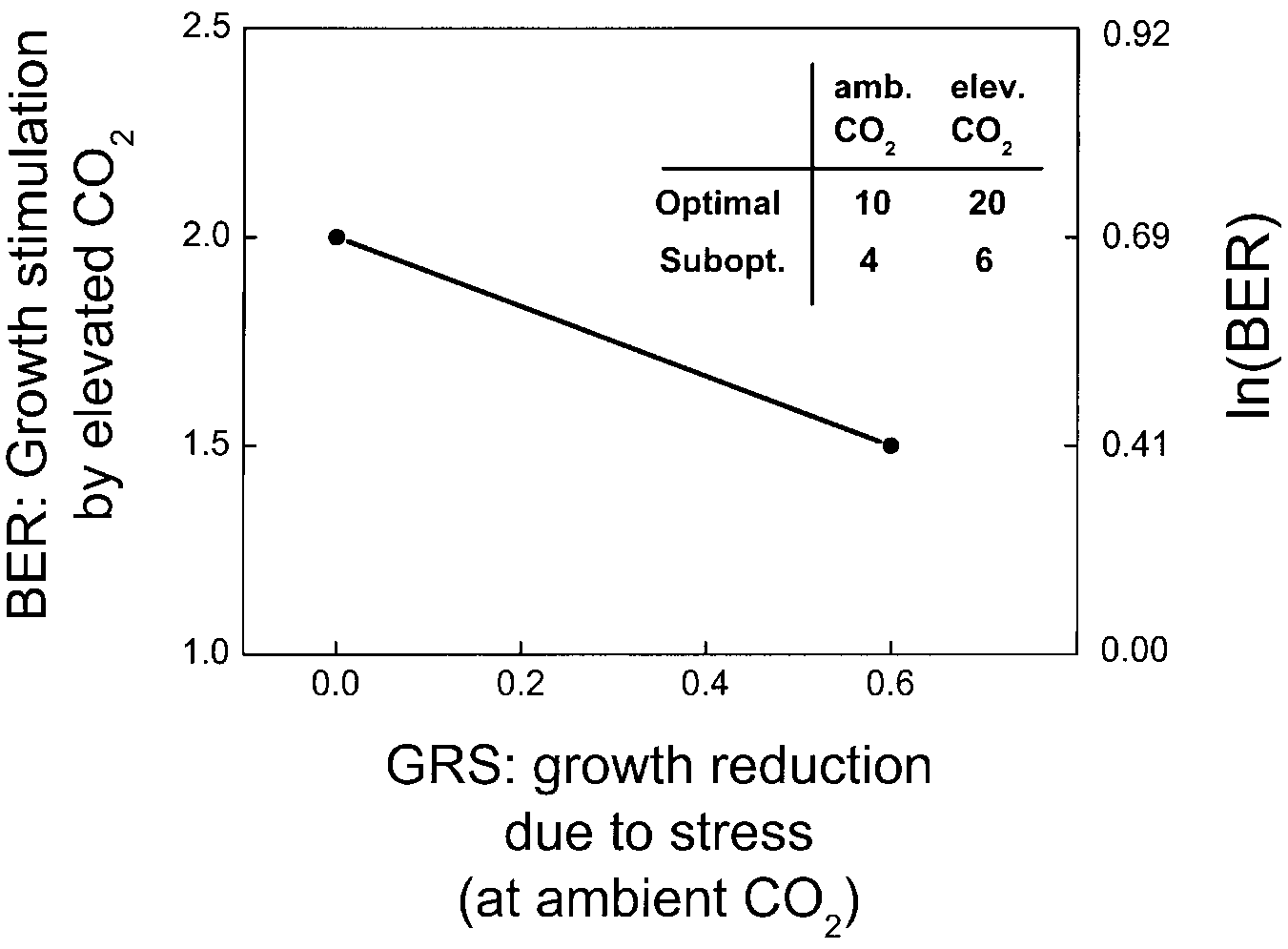
where SLB is an acronym for 'slope of the line connecting the two
the first part of this paper, we analyse the degree of vari-
BER values'. A graphical example of our method is given in
ability in the results of a CO ×environment interaction
Fig. 1. If plant biomass is as specified in the insert, then the ratio
when we repeatedly sample a limited number of plants
of plant biomass at elevated CO relative to ambient CO (BER) is
from the same experimental population.
2 at the optimal level and 1.5 at the sub-optimal level. At ambientCO , we assume that the treatment with the highest biomass is op-
In the second part, we try to obtain an overall picture
timal, with a GRS of 0 as the x-value at which we plot the BER of
of the interaction between elevated CO and environ-
2. The growth reduction due to the sub-optimal level is 0.6, the x-
mental factors, such as primary resources, temperature
value at which we plot the BER of 1.5. These values result in an
and air pollutants. We will restrict our analysis to indi-
SLB of -0.48. A negative SLB indicates that at a given non-opti-mal level of the interacting factor, the relative growth response to
vidually grown plants in the vegetative stage. Apart from
elevated CO is smaller than under optimal conditions. Note that
the stochastic variation mentioned above, another factor
in most of this paper we will focus on the relative growth re-
may hinder generalisations across experiments, i.e. the
sponse; the absolute growth response will almost always be lower
range of environmental growth conditions applied in dif-
A weak point in this approach is that we assume that BER
ferent experiments, which most likely stress plants to
changes linearly from optimal to non-optimal levels and that the
different degrees. Therefore, we follow a method that
environmental condition at which plants show the largest growth
links the growth stimulation due to elevated CO to the
growth reduction at ambient levels of CO due to the
stress factor. That is, the severity of the applied environ-mental stress, as evident from the growth reduction inthe control CO plants, is used to scale the change in bio-
mass response to elevated CO. This allows one to, at
least partly, correct for differences between experiments. An additional advantage of this approach is that we cancompare interactions between elevated CO and a range
of growth-limiting environmental factors at the samescale.
SLB, a parameter to quantify CO ×environment interactions
The minimal experimental design to analyse CO ×environment in-
teractions requires an orthogonal combination of two CO concen-
trations (ambient and elevated) and two levels of the other envi-
Fig. 1 Example to show the method used to calculate the effect of
ronmental factor (optimal and non-optimal for growth). To quanti-
limiting factors on the biomass enhancement ratio (BER). The x-
tatively analyse those experiments, we used a method based on
axis represents the reduction in total biomass at ambient CO of
two main parameters. The first is an indicator of the stimulating
plants grown at the sub- or supra-optimal level when compared to
effect of elevated CO on total plant biomass (sum of above- and
the total biomass of plants grown at the optimal level (growth re-
belowground biomass) and is calculated as the ratio of plant bio-
duction due to stress, GRS). The y-axis represents the ratio of
mass at elevated and at ambient CO levels. We call this the 'bio-
plant biomass at elevated and ambient CO levels. The positive,
mass enhancement ratio', using BER as an acronym. The second
zero or negative sign of the slope of the line connecting the two
parameter is an indicator of the stress experienced by plants due to
BER values indicates the type of interaction (see text). For the cal-
a non-optimal level of the environmental factor under study. For
culations, all BER values have to be ln-transformed prior to any
each experiment, we considered as the 'optimal' level, the treat-
statistical analysis, as ratios are ln-normally distributed by nature.
ment that resulted in the highest total biomass. The intensity of the
treatments. The slope calculated to determine a CO ×environment
interaction is based on the biomass of at least four differentlytreated groups of plants, each with its own variability in total bio-mass. Consequently, the estimate of the slope is affected by theadded variability in all four experimental groups (cf. Poorter et al. 1996; Hedges et al. 1999; Jasienski and Bazzaz 1999). The preci-sion of the slope is co-determined by the number of plants harvest-ed per treatment. Because of constraints on space and labour, thenumber of replicates harvested per treatment in experiments thatstudy CO effects in combination with other factors will generally
be low. This is unfortunate, because it decreases precision where-as, in fact, due to the added variability in four plant groups, ahigher number of replicates would have been required than in asingle-factor experiment with two groups of plants.
To what extent might plant-to-plant variability explain the ob-
served variation in SLBs as in Fig. 2? Because we do not know allthe details of each experiment, we can only answer this questionby a simulation of the most likely situation. From the specifica-tions of the CO ×nutrient experiments provided by the authors, we
know that the median number of plants harvested per treatmentwas five. A low number is four, and a high number is ten, asjudged from the 20th and 80th percentile, respectively, of the com-
Fig. 2 Frequency distribution of CO ×nutrient interactions. Bars
piled number of plants harvested in these experiments. We do not
indicate SLB values derived from 123 published observations. The
know the variability in the plant populations under investigation. bold line indicates the distribution of SLBs after simulating a
Poorter and Garnier (1996) used the standard deviation in ln-trans-
range of experiments with a low (n=4), an intermediate (n=5) and
) as a way to characterise variability in ex-
a high (n=10) number of replicates per treatment, harvesting plant
perimental plant populations. From their compilation of a range of
=0.51) variability in dry mass. The aver-
low value of 0.21 (20th percentile) and a high value of 0.51 (80th
age mass for the four different treatments was chosen so that both
percentile). Assuming now that the true GRS and SLB values
GRS and SLB were exactly the same as the average values in the
were the average of the 123 experimental observations (0.55 and
compiled data set. More information is given in the text
-0.41, respectively), and that plant-to-plant variability is not al-tered by elevated CO , we simulated experiments in which we ran-
domly 'harvested' four, five or ten plants out of three artificial
response is truly optimal; this may not necessarily be the case. An
of 0.21, 0.31 or 0.51, respectively. In this
advantage is that the same method can be applied to different en-
way, we arrived at nine different scenarios, and for each of these
vironmental variables, since the interactive effect with elevated
CO is related to the growth reduction caused by the non-optimal
assume that the aggregated distribution of calculated slopes gives
level and not to the environmental level itself. This enables a com-
us a reasonable estimate of the extent to which slopes vary due to
parison of different treatments, using the growth reduction due to
random variation in biomass alone. The simulated distribution of
the stress factor as a biological yardstick.
SLB values is shown as the continuous line in Fig. 2. Although the
Biomass responses were analysed based on a compilation of
'true' (average) SLB value was negative, positive interactions
published and unpublished experiments on individually grown
were observed in 22% of the simulations. Moreover, variation was
herbaceous and woody C species (see Appendix 1 and 2). C spe-
largely similar to that observed in the literature. Based on this sim-
cies were excluded, because the low number of CO ×environment
ulation, we conclude that the relatively low number of plants har-
studies conducted with these plants hardly allows any generalisat-
vested from rather variable populations can explain most of the
ion. In addition, we did not consider those studies in which the
observed variability in CO ×nutrient interactions. We do not doubt
non-optimal treatment caused a growth reduction of less than
that variation in SLB is also partly due to differences between spe-
10%, both because we felt that such a treatment was not stressful
cies or growth conditions. However, in our opinion, support for
for the plants and because the GRS would become too small to ac-
these alternative explanations has to be found in an a posteriori
curately determine the slope of the line in Eq. 2. Following the
analysis of a range of experiments and not in the mere observation
above method, we calculated the SLBs for a range of factorial ex-
that species A in experiment 1 responded differently from species
periments, restricting the analysis to plants in the vegetative phase.
B in experiment 2 (see also General discussion below). In the
The ambient CO concentration ranged between 300 and 400 µl l-1,
analysis to follow, we will consider the average response across all
and the elevated CO concentration between 550 and 1,100 µl l-1,
observations, and only test for possible differences between herba-
except for one experiment with high salinity.
ceous and woody species in general, unless otherwise stated.
How precisely can an interaction be determined?
The SLB values may differ substantially between experiments. Anexample is given in Fig. 2, where we plotted the distribution of
SLB values for 123 observations of plant species grown in a facto-rial combination of elevated CO and nutrient supply (grey bars).
In some cases, strong positive interactions were reported (e.g.
From the literature data listed in Appendix 1 and 2 and
Whitehead et al. 1997: SLB>1); in other cases, strong negative
plotted in Fig. 2, we obtained the distribution of the
SLBs were found (e.g. Heath and Kerstiens 1997: SLB<-2). Most
slopes represented by the boxplots of Fig. 3. On average,
discussions almost automatically assume that such contrasting re-
the SLB for nutrients was negative (P<0.001), with no
sponses are due to the fact that different experiments use different
indication of a difference between herbaceous and
species, another level of the stress factor, or simply a differentcombination of growth conditions (e.g. Lloyd and Farquhar 2000).
woody species (P>0.5). This implies that a decrease in
A factor that has received less attention is plant variability within
nutrient availability reduces the relative growth response
Theoretically, the relative stimulation of photosynthesisby elevated CO is strongest close to the light compensa-
tion point (Kimball 1986a), and this has indeed been ob-served (Idso and Idso 1994). At low light, plant growthis strongly carbon limited, and therefore one would ex-pect this stimulation of photosynthesis by elevated CO2
to be translated into increased growth. However, analysisof the limited information (Fig. 3; 19 observations)shows that this interaction is small: the average SLBdoes not deviate significantly from zero, although itcomes close (0.05<P<0.1). Similar results have beenfound for crop yield (Kimball 1986a). Although not sig-nificant (P>0.3), there seems to be a tendency for tree
Fig. 3 Distribution of slopes (SLB), indicating the strength of the interaction between elevated CO and the primary resources (nu-
seedlings to have positive SLB values, whereas the her-
trients, irradiance and water) on plant growth. For each of the en-
baceous plants in our compilation showed - on average -
vironmental factors, data are separated for herbaceous species
no response. One might expect tree seedlings to be gen-
(open boxplots) and tree seedlings (shaded boxplots). Data are
erally more shade-tolerant than the five crop species that
based on a literature review of factorial experiments with combi-nations of elevated CO and nutrients (n=51 and n=72 for herba-
represent the herbaceous plants in this case. Such obser-
ceous and woody species, respectively, in 83 papers), irradiance
vations would be in line with the conclusion of Kerstiens
(n=11 and n=8, respectively, in 8 papers) and water (n=12 and
(1998) that within the group of woody species, the
n=30, respectively, in 25 papers). An explanation of SLB values is
shade-tolerant ones are the strongest in their growth re-
given in Methodology and Fig. 1. Numbers in the graph are the
sponse. He suggests that shade-tolerant species have a
10%-trimmed means of SLB values for herbs and woody speciestogether. Boxplots indicate the distribution of a range of observa-
lower leaf area per unit leaf mass, which is less reduced
tions. The lower part of the box shows the 25th percentile. The
than in other tree species at elevated CO. In addition,
highest part of the box gives the 75th percentile, and the line in
species-specific differences in response in tree seedlings
between, the median (50th percentile). The whiskers indicate the
may change with small increases in light availability
10th (lower) and 90th (higher) percentile
(Hättenschwiler and Körner 2000). Clearly, the numberof experiments with low light is far too limited to allow
of plants to elevated CO. Similar conclusions have been
any firm conclusion. Moreover, other factors like the
drawn for CO -enriched crops (Kimball 1986a) and veg-
quality of light used in the experiments may play a role
etations (Stöcklin et al. 1998). Overall, the pattern of re-
sponse was not affected by the type of nutrient in shortsupply, as judged from the similarity in interaction be-tween experiments where nitrogen, phosphorus or all nu-
trients together were modified (Poorter 1998). Althoughthe average SLB is negative, positive slopes are found in
Overall, the results obtained for a range of different her-
20% of the experiments. As discussed below, more de-
baceous and woody species confirmed that a reduced
tailed research, including a range of nutrient levels,
water supply modestly enhances the relative growth re-
should show whether these positive slopes are merely
caused by chance or are a systematic response of specific
P<0.05), with again a small but non-significant differ-
ence between herbs and trees (0.05<P<0.1). As in the
At low nutrient levels, growth is apparently not re-
case of nutrients, 20% of the observations show an inter-
stricted by carbon availability, since high concentrations
action deviating from the general trend.
of starch and other non-structural carbohydrates are usu-
ally found in nutrient-limited plants. Therefore, we do
30-60% on average (Morison 1993), which in turn re-
not expect an increase in carbon fixation to lead to a sim-
duces water loss in the plant. Consequently, CO may al-
ilar stimulation in growth, unless plants at elevated CO
leviate plant water stress by reducing water use. Howev-
would acquire more nutrients or use them more efficient-
er, plants that are stimulated in growth by high CO will
ly (BassiriRad et al. 2001). In the case of N, one of the
have an increased leaf area. This will result in increased
ways to use the acquired nutrients more efficiently is to
transpiration at the whole-plant level, thereby moderat-
invest less of the available N into Rubisco, and more into
ing the interaction (Samarakoon and Gifford 1996). The
other compounds that limit growth. Interestingly, this
effect of CO on stomatal conductance is observed in
does not happen (Medlyn 1996; Makino et al. 2000). We
are only beginning to understand the mechanism by
throughout plant development, with little evidence for
which plants with a low nutrient status adjust their
acclimation. There is growing experimental evidence
growth and how this limits the response to elevated CO
suggesting that elevated CO may have small or insignif-
icant effects on stomatal conductance of many forest tree
species, especially conifers (Curtis 1996). Hence, the re-duced use of water in coniferous forests growing underelevated CO and the subsequent growth response may
be smaller than predicted. In our compilation, however,we did not find a difference in the strength of the interac-tion between conifers and hardwoods (P>0.7).
Interaction with temperature and salinity
Our analysis shows that the average SLB is negative forsub-optimal temperatures, which indicates that at close-to-optimal temperatures, the relative biomass increase byelevated CO is higher than at low temperatures (Fig. 4;
59 observations, P<0.001). This result is in agreementwith results from previous analyses, which also conclud-
Fig. 4 Distribution of SLB values, indicating the strength of the interaction between elevated CO and sub-optimal temperature,
ed that low temperature reduced the growth response to
supra-optimal temperature and salinity. Data are based on a litera-
elevated CO (Idso et al. 1987; Rawson 1992; Curtis and
ture review (sub-optimal temperature: n=48 and n=11 for herba-
Wang 1998), although, again, 20% of the observations
ceous and woody species, respectively, in 24 papers; supra-opti-
differ in direction from the other experiments, with a
mal: n=5 and n=4 in 6 papers; salinity: n=16 and n=2 in 12 pa-pers). Because of the low number of observations for supra-opti-
BER higher at low temperature. No statistical difference
mal temperatures and for woody species at high salinity, we only
was detectable between herbs and woody species
calculated the average values (open circles herbaceous plants,
(P>0.15). In a few experiments, the highest temperature
closed circles woody plants). For more information see the legend
was supra-optimal for growth. In those cases, the largest
growth response was at the highest temperature as well,although the difference was not statistically significant(Fig. 4; 9 observations, P>0.15).
negative effects of a supra-optimal salt (NaCl) concen-
There are at least two explanations for the CO ×tem-
tration on growth. This has indeed been found in a num-
perature interaction. In the short term, an increase in am-
ber of cases, but not all, and the mean SLB does not de-
bient CO concentration results in increased photosyn-
viate significantly from zero (Fig. 4; 18 observations,
thesis in C species, not only by increasing the concen-
P>0.4). Hardly any data have been published for woody
tration of substrate but also by suppressing oxygenation
species. Munns et al. (1999) suggested a positive
(Long 1994). An increase in temperature promotes oxy-
CO ×salt interaction at low salinity, but no CO effect at
genation relative to carboxylation through decreases in
high salinity. From the present compilation we conclude
the affinity of the enzyme Rubisco for CO. Moreover,
that most halophytes have a higher BER at supra-optimal
the solubility of CO decreases faster than that of O at
salinity, whereas most glycophytes have a lower BER
high temperature, diminishing the relative abundance of
under these conditions (Appendix 1 and 2). However, the
CO in the chloroplasts (Jordan and Ogren 1984). There-
few observations available preclude any firm conclusion
fore, the stimulating effect of elevated CO on photosyn-
thesis is strongest under warmer conditions. An alterna-tive explanation for the low response at low temperaturesis that growth is more impaired by sub-optimal tempera-
tures than photosynthesis (Körner 1991; Rawson 1992). As in the case of low nutrient supply, this will result in
the accumulation of non-structural carbohydrates. Withsink strength being so crucial for the growth response of
Of all factors considered here, ozone shows the strongest
plants (e.g. Reekie et al. 1998), plants at low temperature
interaction with CO. The slope is positive (Fig. 5; 29
are probably not able to profit much from an increased
observations, P<0.001), and this is true for 95% of the
sugar supply due to elevated CO (Greer et al. 2000).
observations, with no indication of a difference between
woody and herbaceous species (P>0.7). This implies thatCO strongly ameliorates the detrimental effect of ozone.
There is good evidence that in plants in which stomatalconductance is reduced by CO enrichment, O flux into
Salinity has a negative effect on both the water status
the leaf interior is reduced and this contributes to reduc-
and the photosynthetic apparatus of plants (Ball and
ing the injurious impact of O on plant growth and phys-
Munns 1992). As elevated CO has exactly the opposite
iology (Turcsányi et al. 2000). Three major questions re-
effects, one might expect elevated CO to ameliorate the
main with regard to the protection against O damage
generally increases the concentrations of soluble phenol-ic compounds (Poorter et al. 1997; Peñuelas and Estiarte1998), some of which are known to decrease plant sensi-tivity to UV-B. Most results to date have been obtainedunder artificial-environment conditions, which could re-sult in stronger damage than in the field situation. First,the UV-B levels used in the experiments are generallyvery high (Rozema 1993). Second, leaves developed un-der high light adapt morphologically and physiologicallyin a way that may also confer protection against UV-B(Teramura and Murali 1987). Consequently, plants ingrowth chambers, in which the daily irradiance is abouttwo times lower than under field conditions (Garnier andFreijsen 1994), may be more sensitive to UV-B thanplants in the field. Fig. 5 Distribution of SLB values, indicating the intensity of the interaction between elevated CO
based on a literature review of interactions with O (n=16 and
n=13 for herbaceous and woody species, respectively, in 19 pa-
pers), UV-B (n=2 and n=6 in 8 papers) or SO (n=3 and n=0 in 2
papers). Because of the low number of observations for UV-B and
The very few data available on the combined effects of
SO , we only calculated the average values (open circles herba-
elevated CO and supra-optimal SO (3 observations)
ceous plants, closed circles woody plants). For more information
show a positive interaction, with high SLB values. Thissuggests that CO enrichment reduces the adverse effects
of SO on plant growth. SO is probably used as a source
provided by elevated CO. First, does elevated CO in-
of sulphur and assimilated to proteins and other organic
duce other advantageous mechanisms in addition to sto-
matal closure, such as detoxification or repair processes
higher metabolic rates that may stimulate the sulphur as-
(J. Cardoso-Vilhena, personal communication)? Second,
similation and accelerate repair processes (Rao and De
what is the combined effect of elevated CO and O on
Kok 1994). In addition, high CO decreases stomatal
the growth and productivity of species in which the sto-
conductance, which in turn may reduce the SO flux into
mata are less responsive to CO enrichment, such as
the leaf. However, when SO levels are very high, as in
many conifers? Data indicate that for these species, there
many East European countries, elevated CO may not be
may be similar effects of O at ambient and elevated
able to counteract the detrimental effect of SO.
CO , or at least much less amelioration of O damage
than observed in herbaceous species (Pérez-Soba et al. 1995). However, the data on conifers in the literature are
at present too sparse to be conclusive at this stage. Andthird, what is the combined effect of elevated CO and
O on photosynthesis? Long-term exposure to elevated
CO is accompanied by a decrease in Rubisco activity or
We would like to make a strong case for meta-analysis as
amount of Rubisco protein in many species (Drake et al.
a tool that allows generalisation across a wide range of
1997). Likewise, both short-term exposures to peak con-
experiments (Gurevitch and Hedges 1999). It provides a
centrations of O and to high background concentrations
framework to judge whether a new result falls within the
of O show a decline in Rubisco activity (Pell et al.
low, high or average range of previous observations.
1994). If the effects of elevated CO and elevated O on
Moreover, it may allow the detection of contrasting re-
Rubisco were additive, then the decrease in activity
sponses between (groups of) species or environments,
would result in a reduction of photosynthetic capacity.
before such differences have been explicitly tested in aspecifically designed experiment. Finally, because thestrength of the interaction is prone to random variation
(Fig. 2), average values across experiments may give abetter estimate of the strength of the interaction under
Experiments with CO ×UV-B interactions are scarce
study. However, when interpreting the results of a meta-
(8 observations). As with other interactions, data are
analysis, one should keep in mind that this approach has
variable, and the average SLB does not deviate signifi-
some limitations. First, unnoticed mistakes may have oc-
cantly from 0 (Fig. 5; P>0.5). Thus, elevated CO may
curred in the experimental phase or during calculation of
not compensate for the harmful effect of UV-B. The rea-
the data on which the compilation is based. Second, re-
son for this could be that UV-B primarily affects photo-
searchers may have chosen to refrain from publishing
system II, whereas CO influences carboxylation and
data that were found to be statistically non-significant,
stomatal conductance. On the other hand, elevated CO
which may bias the overall picture (Gurevitch and Hedges
1999). Third, the available studies are not necessarily aweighted random sample of global vegetation, implyingthat estimates of the response of the 'average' C plant
or vegetation are extrapolations with unknown confi-dence margins. Fourth, we can never exclude that an ob-served class difference in SLB (e.g. woody plants versusherbs) is confounded with another difference across spe-cies (e.g. sun versus shade species), or a difference in ex-perimental conditions (cf. Lloyd and Farquhar 2000). Such a risk is particularly evident when only a few stud-ies have been carried out, as in the case of CO ×light in-
teractions. A last point to consider, especially in the con-text of the present review, is that we assumed that inter-actions would be similar for CO concentrations ranging
between 550 and 1100 µmol mol-1, and that the BERvalues change linearly between the assumed optimal and
Fig. 6 Summary of the average growth response of plants for an
interaction between elevated CO and other environmental factors.
Responses are calculated using a biomass enhancement ratio of
Given these considerations we face a dilemma. Ideal-
1.47 for plants grown under optimal conditions. The average slope
ly, conclusions would be based on large-scale experi-
was calculated from the data of Figs. 4, 5 and 6, and the average
ments that study CO ×environment interactions for a
reduction in growth at 350 µl l-1 CO as calculated in the compiled
wide range (say >15) of ecologically contrasting species.
literature. The dashed line indicates the biomass enhancement byelevated CO that would compensate for biomass reduction under
Even in this case, true generality is only achieved if re-
stress conditions, not only in a proportional but also in an absolute
searchers at different laboratories independently arrive at
similar conclusions. As such large-scale screenings arerare, and the vast majority of experiments is restricted toone to four species, we have to accept that most of the
(e.g. Idso and Idso 1994) do not hold. The growth en-
generalisations will come from combining information
hancement by elevated CO is severely reduced at low
from a variety of experiments. To minimise the chance
temperatures or poor nutrient supply. This is not only ex-
effect alluded to in Fig. 2, we suggest using an experi-
plained by the more negative SLB values, but also by the
mental design with more than two levels of the interact-
generally strong growth reduction in those experiments
ing factor, giving more degrees of freedom to estimate
(GRS >0.5). The average growth enhancement by elevat-
the overall response. Moreover, if plant-to-plant varia-
ed CO at optimal conditions is not significantly altered
tion is not of prime interest, all precautions possible
by high UV-B, high salinity or low irradiance, mainly
should be taken to minimise and control plant-to-plant
because the average SLB values were only marginally
variability within the experimental population (Poorter
different from zero. The interaction with water was sig-
and Garnier 1996), which will also improve the precision
nificant, but the effect was small. The interaction be-
tween elevated CO and O was strong. This is the only
type of stress where biomass is stimulated more thantwofold under elevated CO (BER values at high O are
often larger than 2). The average value is above the dot-ted line, indicating that the loss of biomass at elevated
The effect of an interaction between CO and any envi-
O is more than compensated by the presence of elevated
ronmental factor will not only depend on the slopes of
CO. However, the biomass of high-CO plants at high
the lines (Figs. 3, 4 and 5), but also on the magnitude of
O concentrations is not as large as that of high-CO
the growth reduction due to the stress factor at ambient
CO. This is taken into account in Fig. 6, where we plot
the average BER values against the average GRS, as ex-plained in Fig. 1. At optimal conditions (GRS=0), we as-
sumed a BER value of 1.47 (average from the compila-tion by Poorter et al. 1996). The BER values at non-opti-
The responses in Fig. 6 are average values of literature
mal conditions were then derived from the average SLB
data for both herbaceous and woody species. Some time
and GRS values in the present compilation. The dashed
ago, Curtis and Wang (1998) reviewed the growth re-
line in the figure indicates the extent to which the en-
sponse of woody plants to elevated CO. To the extent
hancement in plant biomass by elevated CO should in-
crease in order to compensate for growth losses at non-
conclusions and ours are in agreement. This can be ex-
optimal conditions, not only in a proportional but also in
plained by the fact that we did not find systematic differ-
an absolute way. Clearly, propositions that plants under
ences between woody seedlings and herbaceous species
stress will always respond relatively more strongly to
for any of the environmental factors, although some (ir-
enrichment than those under optimal conditions
radiance, water) are on the verge of significance. Con-
clusions deviate strongly for the factor ozone, where we
calculated much stronger responses both for herbaceousand woody species. The fact that Curtis and Wang
Plant-to-plant variability in biomass within treatments is
(1998) had only two data points for this factor may ex-
one of the factors that explains contrasting CO ×environ-
plain the different results. We were not able to find sys-
ment interactions published in the literature. On average,
tematic differences in the compiled literature between re-
the growth stimulation by elevated CO is smaller at low
sponses of gymnosperms and hardwood seedlings. This
nutrient availability and low temperature, increases
may imply that the differential response of stomatal con-
somewhat at low water supply, and is substantially high-
ductance with respect to increased CO does not neces-
er at high ozone concentrations. There is a strong paucity
sarily lead to a strongly different CO ×environment in-
of data on the interaction with light, salt, UV-B, nitroge-
nous air pollutants and SO , but, with the exception of
We have not paid attention to C and Crassulacean ac-
SO , average responses are small. No systematic differ-
id metabolism species, because far less information is
ences were found between woody and herbaceous spe-
available for the response of these species under sub- or
supra-optimal conditions. However, as their response toelevated CO is generally smaller than that of C species
Acknowledgements We thank Ep Heuvelink, Eric Garnier, Gina
(Poorter et al. 1996), we expect the CO ×environment
Adams and Manuela Chaves for trustfully providing us with (par-
tially unpublished) data for incorporation in our analyses. InekeStulen, Jan Goudriaan, Marcel van Oijen and an anonymous re-viewer thoughtfully commented on a previous version of themanuscript.
×environment interaction. Data are for herbaceous species and listed in alphabetical order
ences are given as first author and year of the publication
olin (1996) Physiol Plant 97:674-684; V
illiams (1981) Plant Physiol 68:1406-1409;
ard (1999) Global Change Biol 5:857-867;
fects of climate change on marine coastal ecosystems,
1; Nicolas (1993) Aust J Plant Physiol 20:349-360; Oberbauer
17-129; Reinert (1997) New Phytol 137:41
eed Sci 36:751-757; Radoglou (1992) Ann Bot 70:245-256; Reddy (1998) En-
Crop Sci 38:122-128; Nakamura (1999) Photosynthetica 37:61-72; Newbery (1996)
(1986) Can J Bot 64:2993-2998; Olszyk (1997) Agric Ecosyst Environ 66:1-10; Paez
(1984) J Agric Sci 102:687-693; Patterson (1982) W
(1997) Plant Ecol 128:182-191; Rufty (1994) Physiol Plant 91:503-509; Sa (1998) J.
Plant Nutr 21:2207-2218; Samarakoon (1995) Aust J Plant Physiol 22:33-44; Schwarz
(1984) J Exp Bot 35:193-196; Serraj (1999) Global Change Biol 5:283-291; Sionit
(1981) Agron J 73:1023-1027; Sionit (1982) Agron J 74:721-725; Sionit (1983) Crop
Sci 23:329-333; Sionit (1987) Can J Plant Sci 67:59-67; Sritharan (1992) Garten-
bauwissenschaft 57:246-251; Stirling (1998) New Phytol 140:343-354; T
Wheeler (1991) Crop Sci 31:1209-1213; W
(2000) Plant Physiol 124:767-779; Zerihun (2000) Ann Bot 86:723-730; Ziska (1997)
18-123; Imai (1979) Jpn J Crop Sci 48:409-417; Israel
ageningen, pp 64-67; Lenssen (1993a) PhD thesis, Free
126; Carlson (1982) Oecologia 54:50-54; Chen (1997) Gartenbauwis-
1; Barett (1995) J Biogeogr 22:331-339; Barnes (1992) New Phytol 121:403-412;
field; Almeida (1999) Plant Soil 210:159-166; Arp (1998) Plant Cell Environ
Aben (1999) Aust J Plant Physiol 26:759-766; Adams (1996) PhD thesis, University of
Barnes (1995) Global Change Biol 1:129-142; Bernacci (2000) Global Change Biol
6:855-863; Bowler (1993) New Phytol 144:515-522; Bowler (1996) New Phytol
132:391-401; Campbell (1993) Proc 17th Grassland Conf, Palmerston North, New Zea-
senschaft 62:30-37; Coleman (1992) Ecology 73:1244-1259; Coleman (1993) Oecologia
93:195-200; Cure (1988) Crop Sci 28:671-677; De Luis (1999) Physiol Plant
107:84-89; Ferris (1995) New Phytol 131:491-501; Ghanoum (1998) Aust J Plant
Physiol 25:627-636; Goudriaan (1983) Neth J Agric Sci 31:157-169; Greer (2000) Aust
J Plant Physiol 27:301-310; Harmens (2000) Ann Bot 86:833-839; Heagle (1993) New
Phytol 123:751-762; Heagle (1999) Crop Sci 39:731-744; Hocking (1985) Ann Bot
55:835-844; Hughes (1971) Ann Bot 35:933-945; Hunt (1995) Ann Bot 75:207-216;
(1990) J Plant Nutr 13:1419-1433; Jablonski (1997) Can J Bot 75:533-545; Jansen
(1986) Biological control of photosynthesis, Nijhof
(1996) J Environ Qual 25:908-916; Larigauderie (1988) Oecologia 77:544-549; Lenssen
(1990) In: Goudriaan et al (eds) The greenhouse ef
Oecologia 84 207-214; McConnaughay (1993) Oecologia 94:550-557; Miller (1998)
×environment interaction. Data are for woody species listed in alphabetical order
given as first author and year of the publication
Arnone (1990) New Phytol 116:55-66; Arp (1998) Plant Cell
Garnier E, Freijsen AHJ (1994) On ecological inference from lab-
Environ 21:1-11; Ball (1997) Plant Cell Environ 20:1158-1160;
oratory experiments conducted under optimum conditions. A
Bassow (1994) Ecol Appl 4:593-603; Bazzaz (1993) Ecology
Whole Plant Perspective on Carbon-Nitrogen Interactions.
74:104-114; Broadmeadow (2000) New Phytol 146:437-451;
Roy J, Garnier E (Eds) SPB Academic Publishing, The Hague,
Brown (1986) Tree Physiol 2:223-232; Bruhn (2000) New Phytol
146:415-425; Catovsky (1999) Global Change Biol 5:507-518;
Gifford RM (1992) Interaction of carbon dioxide with growth-lim-
Centritto (1999) New Phytol 141:119-140; Conroy (1986) Ann
iting environmental factors in vegetation productivity: impli-
Bot 57:165-177; Conroy (1988) Plant Cell Environ 11:91-98;
cations for the global carbon cycle. Adv Bioclimatol 1: 24-58
Conroy (1990) Plant Cell Environ 13:329-337; Conroy (1992)
Greer DH, Laing WA, Campbell BD, Halligan EA (2000) The ef-
Plant Cell Environ 15:843-847; Curtis (1995) New Phytol
fect of perturbations in temperature and photon flux density on
129:253-263; Dickson (1998) Can. J For Res 28:1706-1716; El-
the growth and photosynthetic response of five pasture species
Kohen (1992) Ann Sci For 49:83-90; El-Kohen (1994) Tree
to elevated CO. Aust J Plant Physiol 27: 301-310
Physiol 14:679-690; Gaucher (1998) In: De Kok and Stulen (eds)
Gurevitch J, Hedges LV (1999) Statistical issues in ecological me-
Responses of plant metabolism to air polution and global change,
Backhuys, Leiden, pp 305-308; Gebauer (1996) New Phytol
Hättenschwiler S, Körner C (2000) Tree seedling responses to in
134:85-93; Gleadow (1998) Plant Cell Environ 21:12-22;
situ CO -enrichment differ among species and depend on un-
Goudriaan (1983) Neth J Agric Sci 31:157-169; Griffin (1993)
derstorey light availability. Glob Change Biol 6: 213-226
Oecologia 95:575-580; Griffin (1995) New Phytol 129:547-556;
Heath J, Kerstiens G (1997) Effects of elevated CO on leaf gas
Griffin (1997) Plant Soil 190:11-18; Guehl (1994) Tree Physiol
exchange in beech and oak at two levels of nutrient supply:
14:707-724; Heath (1997) Plant Cell Environ 20:57-67; Hibbs
Consequences for sensitivity to drought in beech. Plant Cell
(1995) New Phytol 129:569-577; Hoffmann (2000) Oecologia
123:312-317; Johnsen (1993) Can J For Res 23:1033-1042; John-
Hedges LV, Gurevitch J, Curtis PS (1999) The meta analysis of re-
son (1995) Plant Soil 168/169:535-545; Kerstiens (1994) New
sponse ratios in experimental ecology. Ecology 80: 1150-1156
Phytol 148:607-614; Koike (1993) Proc IGBP Symp 1992, pp
Hodinott J, Scott R (1996) The influence of light and quality and
425-430; Lavola (2000) Physiol Plant 109:260-267; Lewis (1996)
carbon dioxide enrichment on the growth and physiology of
New Phytol 133: 431-443; Lovelock (1996) Funct Ecol
seedlings of three conifer species. I. Growth responses. Can J
10:662-667; Miao (1992) Oecologia 90:300-304; Midgley (1995)
J Biogeogr 22:185-191; Mortensen (1995) Environ Pollut
Idso KE, Idso SB (1994) Plant responses to atmospheric CO en-
87:337-343; Murray (2000) Tree Physiol 20:421-434; Norby
richment in the face of environmental constraints: a review of
(1991) New Phytol 117:515-528; Oberbauer (1986) Can J Bot
the past 10 years' research. Agric For Meteor 69: 153-203
64:2993-2998; Picon (1996) Ann Sci For 53:431-446; Pregitzer
Idso SB, Kimball BA, Anderson MG Mauney JR (1987) Effects of
(1995) New Phytol 129:579-585; Prior (1997) Tree Physiol
atmospheric CO enrichment on plant growth: the interactive
17:397-405; Runion (1999) Tree Physiol 19:329-335; Schorte-
role of air temperature. Agric Ecosys Environ 20: 1-10
meyer (1999) Aust J Plant Physiol 26:737-747; Sheu (1999) Envi-
Jasienski M, Bazzaz FA (1999) The fallacy of ratios and the test-
ron Exp Bot 41:57-65; Silvola (1992) Oecologia 91:208-213;
ability of models in biology. Oikos 84: 321-326
Silvola (1993) Oikos 67:227-234; Silvola (1995) Plant Soil
Jordan DB, Ogren WL (1984). The CO /O specifity of ribulose
168/169:547-553; Stewart (1993) Physiol Plant 88:493-500;
1,5-bisphosphate concentration, pH and temperature. Planta
Sullivan (1994) Plant Cell Environ 17:311-317; Syvertsen (1999)
Plant Soil 208:209-219; Tjoelker (1998) New Phytol
Kerstiens G (1998) Shade-tolerance as a predictor of responses to
140:197-210; Tolley (1984a) Can J For Res 14:343-350; Tolley
elevated CO in trees. Physiol Plant 102: 472-480
(1984b) Can J Bot 62:2135-2139; Townend (1993) Tree Physiol
Kimball BA (1986a) Influence of elevated CO on crop yield. In:
13:389-400; Townend (1995) New Phytol 130:193-206; Tschap-
Enoch HZ, Kimball BA (eds) Carbon Dioxide Enrichment of
linski (1993) Tree Physiol 13:283-296; Tschaplinski (1995) New
Greenhouse Crops Vol. II. Physiology, Yield and Economics.
Phytol 129:63-71; Uselman (2000) Plant Soil 222:191-202; Utri-
ainen (1998) In: De Kok and Stulen (eds) Responses of plant me-
Kimball BA (1986b) CO stimulation of growth and yield under
tabolism to air polution and global change, Backhuys, Leiden, pp
environmental restraints. In: Enoch HZ, Kimball BA (eds)
467-469; Vivin (1997) Ann Sci For 54:597-610; Volin (1996)
Carbon Dioxide Enrichment of Greenhouse Crops Vol. II.
Physiol Plant 97:674-684; Volin (1998) New Phytol 138:315-325;
Physiology, Yield and Economics. CRC press, Boca Raton, pp
Wayne (1998) Oecologia 114:335-342; Whitehead (1997) New
Phytol 135:201-212; Wilkins (1994) Tree Physiol 14:769-779;
Körner C (1991) Some often overlooked plant characteristics as
Wong (1992) Aust J Bot 40:457-472; Yakimchuk (1993) Can J
determinants of plant growth: a reconsideration. Funct Ecol 5:
For Res 24:1-8; Zak (2000) Ecol Appl 10:34-46
Lloyd J, Farquhar GD (1996) The CO dependence of photosyn-
thesis, plant growth responses to elevated atmospheric CO2
concentrations and their interaction with soil nutrient status. I.
General principles and forest ecosystems. Funct Ecol 10: 4-32
Lloyd J, Farquhar GD (2000) Do slow-growing species and nutri-
Ball MC, Munns R (1992) Plant responses to salinity under elevat-
ent-stressed plants consistently respond less to elevated CO ?
ed atmospheric concentrations of CO. Aust J Bot 40:
A clarification of some issues raised by Poorter (1998). Glob.
BassiriRad H, Gutschick VP, Lussenhop J (2001) Root system ad-
Long SP (1994) The potential effects of concurrent increases in
justments: regulation of plant nutrient uptake and growth re-
temperature, CO and O on net photosynthesis, as mediated
sponses to elevated CO. Oecologia 126: 305-320
by RubisCO. In: Alscher RG, Wellburn AR (eds) Plant Re-
Curtis PS (1996) A meta-analysis of leaf gas exchange and nitro-
sponses to the Gaseous Environment. Chapman & Hall, Lon-
gen in trees grown under elevated carbon dioxide. Plant Cell
Luo Y, Mooney HA (1999) Carbon Dioxide and Environmental
Curtis PS, Wang X (1998) A meta-analysis of elevated CO effects
on woody plant mass, form, and physiology. Oecologia 113:
Makino A, Nakano H, Mae T, Shimada T, Yamamoto N (2000)
Photosynthesis, plant growth and N-allocation in transgenic
Drake BG, Gonzalez-Meler MA, Long SP (1997). More efficient
rice plants with decreased Rubisco under CO enrichment. J
plants: a consequence of elevated carbon dioxide? Annu Rev
Plant Physiol Plant Molec Biol 48: 607-640
Medlyn BE (1996) The optimal allocation of nitrogen within the
Rao MV, De Kok LJ (1994) Interactive effects of high CO and
C3 photosynthetic system at elevated CO. Aust J Plant
SO on growth and anti-oxidant levels in wheat. Phyton- Ann
Morison JIL (1993) Responses of plants to CO under water limit-
Rawson HM (1992) Plant response to temperature under condi-
ed conditions. In: Rozema J, Lambers H, van de Geijn SC,
tions of elevated CO. Aust J Bot 40: 473-490
Cambridge ML (eds) CO and the Biosphere. Kluwer Aca-
Reekie EG, MacDougall G, Wong I, Hicklenton PR (1998) Effect
demic Publishers, Dordrecht, pp 193-209
of sink size on the growth response to elevated atmospheric
Munns R, Cramer GR, Ball MC (1999) Interactions between ris-
CO within the genus Brassica. Can J Bot 76: 829-835
ing CO , soil salinity and plant growth. In: Luo Y, Mooney
Rozema J (1993) Plant reponses to atmospheric carbon dioxide
HA (eds) Carbon Dioxide and Environmental Stress. Academ-
enrichment: interactions with some soil and atmospheric con-
Pell EJ, Eckardt NA, Glick RE (1994) Biochemical and molecular
Samarakoon AB, Gifford RM (1996) Water use and growth of cot-
basis for impairment of photosynthetic potential. Photosynth
ton in response to elevated CO in wet and drying soil. Aust J
Peñuelas J, Estiarte M (1998) Can elevated CO affect secondary
Saxe H, Ellsworth DS, Heath J (1998) Tree and forest functioning
metabolism and ecosystem function? Trends Ecol Evol 13:
in an enriched CO atmosphere. New Phytol 139: 395-436
Stitt M, Krapp A (1999) The interaction between elevated carbon
Pérez-Soba M, Dueck TA, Puppi P, Kuiper PJC (1995) Interac-
dioxide and nitrogen nutrition: the physiological and molecu-
tions of elevated CO , NH and O on mycorrhizal infection,
lar background. Plant Cell Environ 22: 583-622
gas exchange and N metabolism in saplings of Scots pine.
Stöcklin J, Schweizer K, Körner C (1998) Efects of elevated CO2
and phosphorus addition on productivity and community com-
Poorter H (1998) Do slow-growing species and nutrient-stressed
position of intact monoliths from calcareous grassland. Oeco-
plants respond relatively strongly to elevated CO ? Glob
Teramura AH, Murali NS. (1987) Intraspecific differences in
Poorter H, Garnier E (1996) Plant growth analysis: evaluation of
growth and yield of soybean exposed to ultraviolet-B radiation
experimental design and computational methods. J Exp Bot
under greenhouse and field conditions. Environ Exp Bot 26:
Poorter H, Roumet C, Campbell BD (1996) Interspecific variation
Turcsányi E, Cardoso-Vilhena J, Daymond J., Gillespie C, Balaguer
in the growth response of plants to elevated CO : A search for
L., Ollerenshaw JH, Barnes JD (2000). Impacts of tropospheric
functional types. In: Körner C, Bazzaz FA (eds) Carbon Diox-
ozone: past, present and likely future. In: Trace Gas Emissions
ide, Populations, Communities. Academic Press, San Diego,
and Plants (ed. by S.N. Singh), Kluwer Academic Publishers,
Poorter H, Van Berkel Y, Baxter B, Bel M, Den Hertog J, Dijkstra
Whitehead SJ, Caporn SJM Press MC (1997) Effects of elevated
P, Gifford RM, Griffin KL, Roumet C, Wong SC (1997) The
CO , nitrogen and phosphorus on the growth and photosynthe-
effect of elevated CO on the chemical composition and con-
sis of two upland perennials: Calluna vulgaris and Pteridium
struction costs of leaves of 27 C species. Plant Cell Environ
aquilinium. New Phytol 135: 201-211
Emerging issues in pharmaceutical and healthcare law and practice Emerging issues in pharmaceutical and healthcare law and practice ACTAVIS v MERCK Ch D (Patents Court) (Warren J) 06/06/2007 The judgment in Actavis v Merck illustrates that in order to obtain patent protection for a second medical use of a known substance, the second use in question must meet the usual pa
Cipro offre una nuova alternativa dove imparare l'Inglese e permette di combinare l'apprendimento della lingua con una vacanza in una località incantevole dove splende sempre il sole. L'isola ha una storia di circa 10.000 anni ed è oggi una modena isola dove la cultura europea e gli splendori di una civiltà antichissima si unioscono. Le spiagge sono meravigliose ed anche il paesaggio intorno
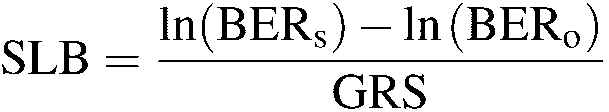
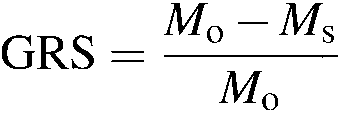
 parameters, such as the distribution and intensity of
stress was then calculated as the reduction in total biomass at am-
clouds (light) and precipitation (water). We therefore
bient CO of plants grown at the non-optimal level compared to
the total biomass of plants grown at the optimal level. We call this
need to analyse how these changing environmental fac-
the 'growth reduction due to stress' (GRS) and calculated it as:
tors may modify the impact of elevated CO on plant
A range of research papers and reviews has dealt with
the interactions between elevated atmospheric CO con-
with M and M being the total biomass of plants at the optimal
level O and at a certain sub- or supra-optimal level S, respectively.
parameters, such as the distribution and intensity of
stress was then calculated as the reduction in total biomass at am-
clouds (light) and precipitation (water). We therefore
bient CO of plants grown at the non-optimal level compared to
the total biomass of plants grown at the optimal level. We call this
need to analyse how these changing environmental fac-
the 'growth reduction due to stress' (GRS) and calculated it as:
tors may modify the impact of elevated CO on plant
A range of research papers and reviews has dealt with
the interactions between elevated atmospheric CO con-
with M and M being the total biomass of plants at the optimal
level O and at a certain sub- or supra-optimal level S, respectively.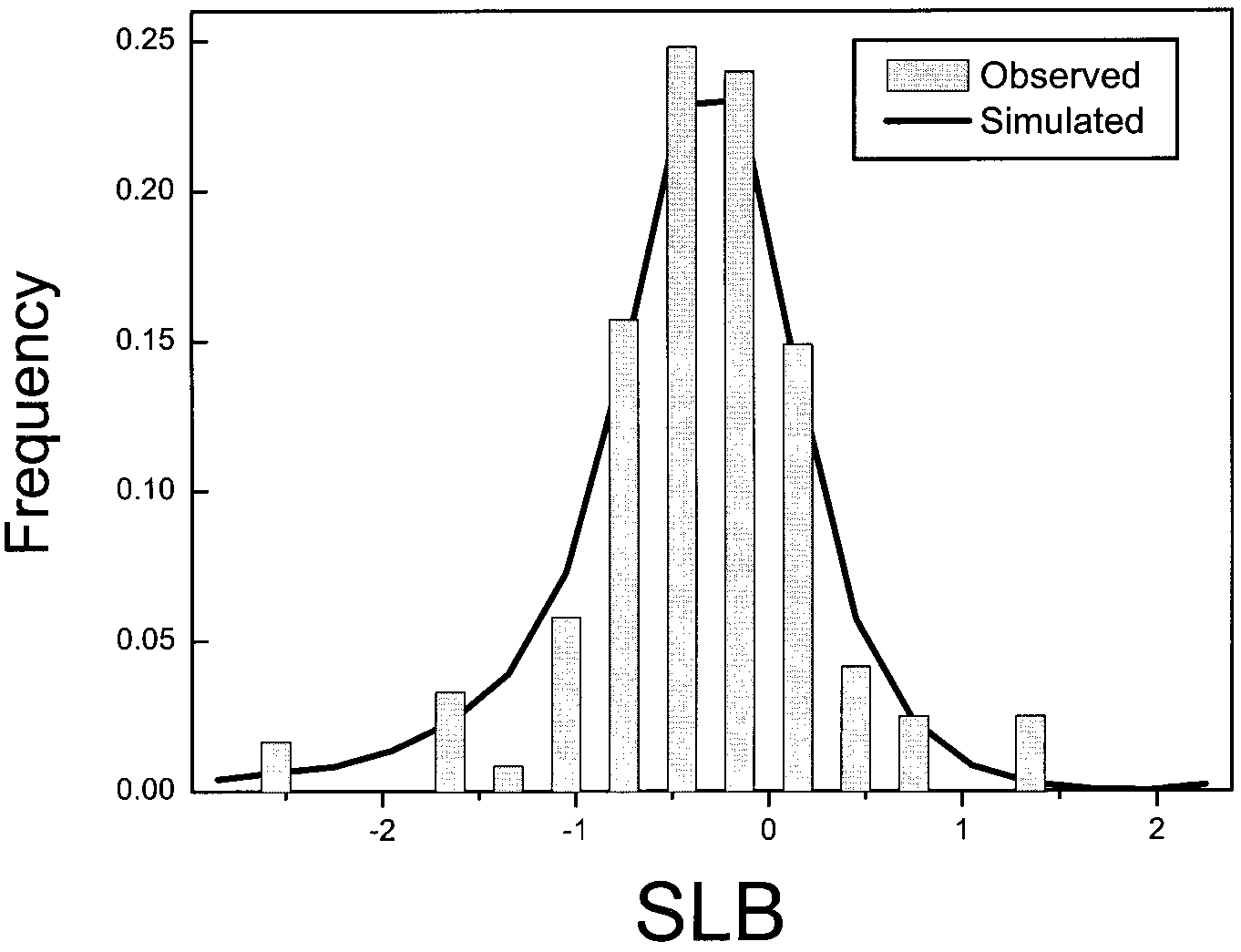 treatments. The slope calculated to determine a CO ×environment
interaction is based on the biomass of at least four differentlytreated groups of plants, each with its own variability in total bio-mass. Consequently, the estimate of the slope is affected by theadded variability in all four experimental groups (cf. Poorter et al.
treatments. The slope calculated to determine a CO ×environment
interaction is based on the biomass of at least four differentlytreated groups of plants, each with its own variability in total bio-mass. Consequently, the estimate of the slope is affected by theadded variability in all four experimental groups (cf. Poorter et al.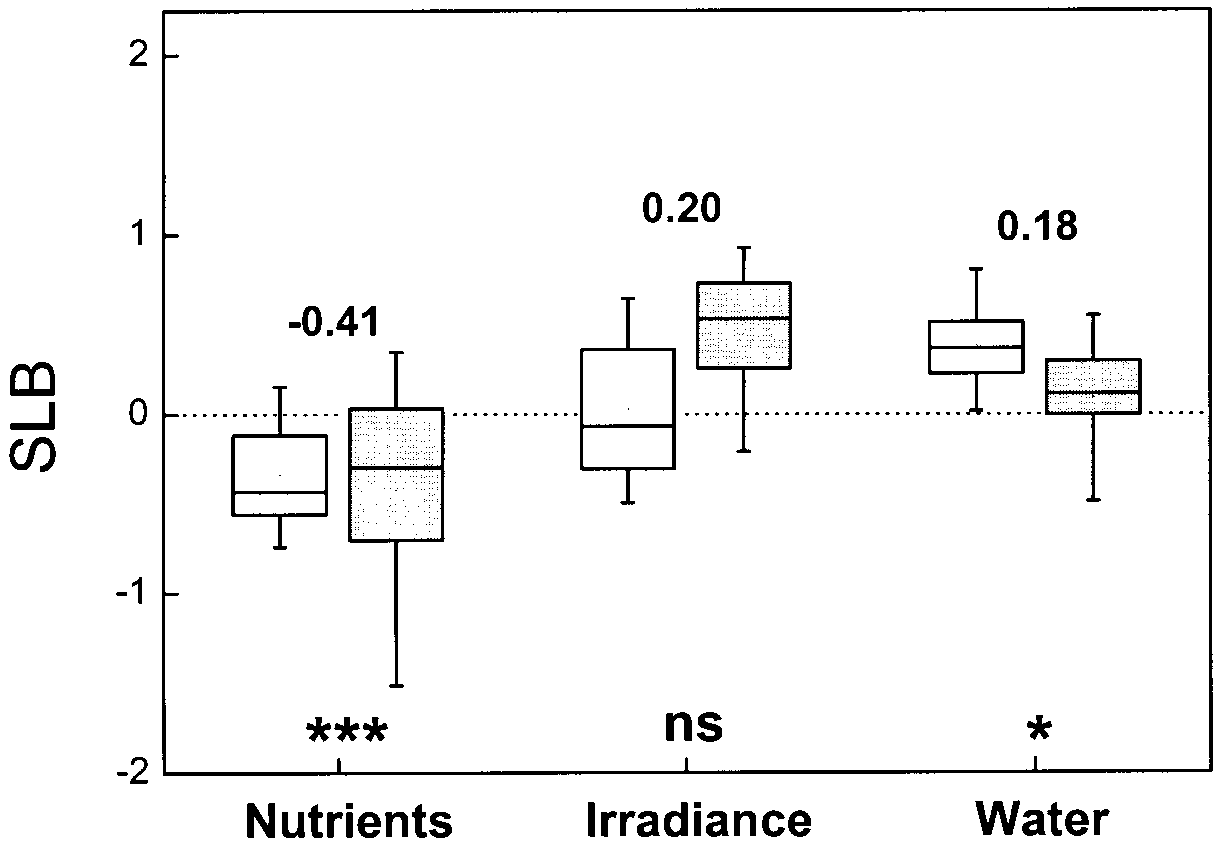 Theoretically, the relative stimulation of photosynthesisby elevated CO is strongest close to the light compensa-
tion point (Kimball 1986a), and this has indeed been ob-served (Idso and Idso 1994). At low light, plant growthis strongly carbon limited, and therefore one would ex-pect this stimulation of photosynthesis by elevated CO2
to be translated into increased growth. However, analysisof the limited information (Fig. 3; 19 observations)shows that this interaction is small: the average SLBdoes not deviate significantly from zero, although itcomes close (0.05<P<0.1). Similar results have beenfound for crop yield (Kimball 1986a). Although not sig-nificant (P>0.3), there seems to be a tendency for tree
Fig. 3 Distribution of slopes (SLB), indicating the strength of the
Theoretically, the relative stimulation of photosynthesisby elevated CO is strongest close to the light compensa-
tion point (Kimball 1986a), and this has indeed been ob-served (Idso and Idso 1994). At low light, plant growthis strongly carbon limited, and therefore one would ex-pect this stimulation of photosynthesis by elevated CO2
to be translated into increased growth. However, analysisof the limited information (Fig. 3; 19 observations)shows that this interaction is small: the average SLBdoes not deviate significantly from zero, although itcomes close (0.05<P<0.1). Similar results have beenfound for crop yield (Kimball 1986a). Although not sig-nificant (P>0.3), there seems to be a tendency for tree
Fig. 3 Distribution of slopes (SLB), indicating the strength of the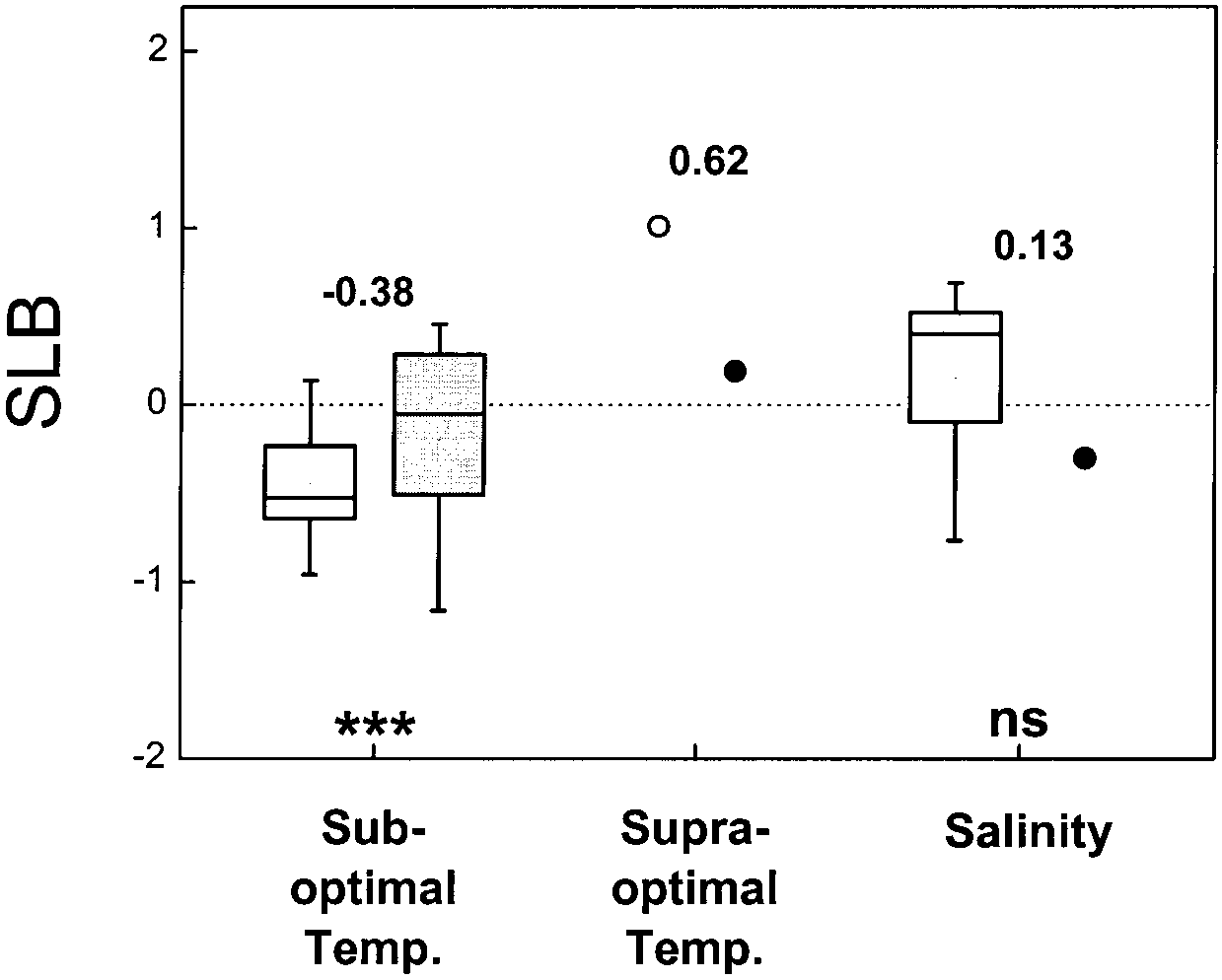 species, especially conifers (Curtis 1996). Hence, the re-duced use of water in coniferous forests growing underelevated CO and the subsequent growth response may
be smaller than predicted. In our compilation, however,we did not find a difference in the strength of the interac-tion between conifers and hardwoods (P>0.7).
species, especially conifers (Curtis 1996). Hence, the re-duced use of water in coniferous forests growing underelevated CO and the subsequent growth response may
be smaller than predicted. In our compilation, however,we did not find a difference in the strength of the interac-tion between conifers and hardwoods (P>0.7).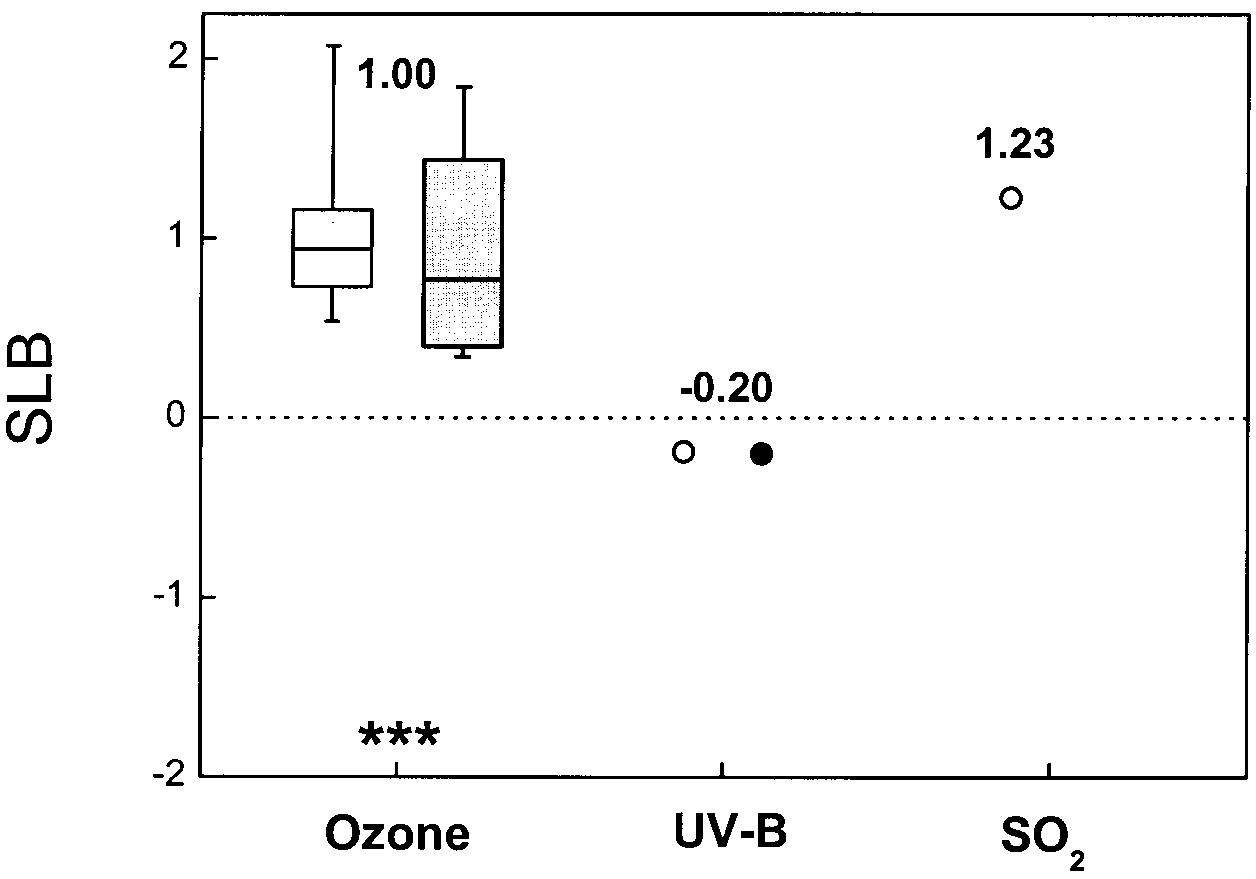 generally increases the concentrations of soluble phenol-ic compounds (Poorter et al. 1997; Peñuelas and Estiarte1998), some of which are known to decrease plant sensi-tivity to UV-B. Most results to date have been obtainedunder artificial-environment conditions, which could re-sult in stronger damage than in the field situation. First,the UV-B levels used in the experiments are generallyvery high (Rozema 1993). Second, leaves developed un-der high light adapt morphologically and physiologicallyin a way that may also confer protection against UV-B(Teramura and Murali 1987). Consequently, plants ingrowth chambers, in which the daily irradiance is abouttwo times lower than under field conditions (Garnier andFreijsen 1994), may be more sensitive to UV-B thanplants in the field.
generally increases the concentrations of soluble phenol-ic compounds (Poorter et al. 1997; Peñuelas and Estiarte1998), some of which are known to decrease plant sensi-tivity to UV-B. Most results to date have been obtainedunder artificial-environment conditions, which could re-sult in stronger damage than in the field situation. First,the UV-B levels used in the experiments are generallyvery high (Rozema 1993). Second, leaves developed un-der high light adapt morphologically and physiologicallyin a way that may also confer protection against UV-B(Teramura and Murali 1987). Consequently, plants ingrowth chambers, in which the daily irradiance is abouttwo times lower than under field conditions (Garnier andFreijsen 1994), may be more sensitive to UV-B thanplants in the field.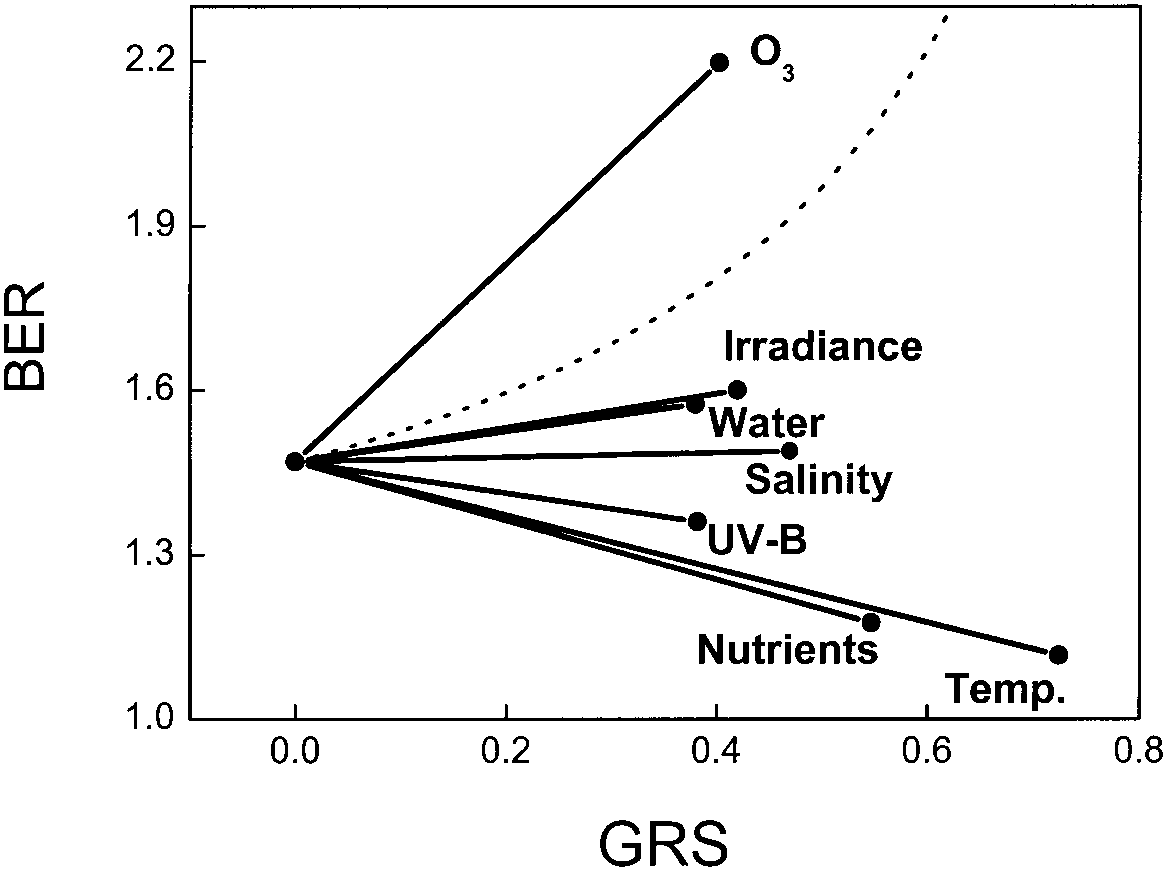 1999). Third, the available studies are not necessarily aweighted random sample of global vegetation, implyingthat estimates of the response of the 'average' C plant
or vegetation are extrapolations with unknown confi-dence margins. Fourth, we can never exclude that an ob-served class difference in SLB (e.g. woody plants versusherbs) is confounded with another difference across spe-cies (e.g. sun versus shade species), or a difference in ex-perimental conditions (cf. Lloyd and Farquhar 2000).
1999). Third, the available studies are not necessarily aweighted random sample of global vegetation, implyingthat estimates of the response of the 'average' C plant
or vegetation are extrapolations with unknown confi-dence margins. Fourth, we can never exclude that an ob-served class difference in SLB (e.g. woody plants versusherbs) is confounded with another difference across spe-cies (e.g. sun versus shade species), or a difference in ex-perimental conditions (cf. Lloyd and Farquhar 2000).