L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
Dyeplants.de
IOBC wprs Bulletin Vol. 27 (3) 2004 European corn borer (Ostrinia nubilalis): Studies on proteinase activity and proteolytical processing of the B.t.-toxin Cry1Ab in transgenic corn
Renate Kaiser-Alexnat, Wolfgang Wagner, Gustav-Adolf Langenbruch, Regina G. Kleespies, Brigitte Keller, Bernd Hommel1 Federal Biological Research Centre for Agriculture and Forestry (BBA), Institute for Biological Control, Heinrichstr. 243, 64287 Darmstadt, Germany (E-mail: R.Kaiser- Alexnat@bba.de); 1BBA, Institute for Integrated Plant Protection, Stahnsdorfer Damm 81, 14532 Kleinmachnow, Germany
Abstract: One possibility to control the European corn borer (ECB) is the cultivation of B.t.-corn. However, this can result in the development of resistant pest populations. To analyse possible mecha- nisms of resistance, a reference system for the identification and quantification of physiological changes in the midgut was established. Studies on proteinase activities were conducted with a suscep- tible German ECB population. The digestive proteinases trypsin, chymotrypsin, elastase, and ami- nopeptidase were identified in the midgut sap of 5th instar larvae. In whole 1st and 2nd instars as well as in the midgut epithelium of 5th instar larvae, the proteinase aminopeptidase was provable. Besides, proteolytical processing of the B.t.-toxin (and protoxin) Cry1Ab as present in transgenic corn is described. Key words : European corn borer, Ostrinia nubilalis, B.t.-corn, midgut proteinases, trypsin, chymotrypsin, elastase, aminopeptidase, B.t.-toxin Cry1Ab, protoxin, proteolytical processing Introduction
In Europe, the economical most important pest in maize (Zea mays L.) is the European corn borer (ECB, Ostrinia nubilalis). Thus, transgenic corn (B.t.-corn) highly insecticidal to the larvae of ECB was developed based on a truncated Cry1Ab toxin of Bacillus thuringiensis. However, the cultivation of the respective cultivars may result in the development of resistant pest populations.
Depending on the mode of action of B.t.-toxins the potential of insect resistance to B.t.-
toxins is generally located at any step of the toxic pathway: ingestion, solubilization, proteolytic processing, binding to specific receptors, membrane integration, pore formation, cell lysis, and insect death (Ferré & van Rie, 2002). Two main mechanisms of resistance to B.t.-toxins have been identified in other pest-B.t.-toxin-systems. One of them is proteinase-mediated and the other receptor-mediated (Oppert et al., 1997, McGaughey & Oppert, 1998).
In order to establish preliminary reference systems for the characterization of potential
available resistant individuals, first studies on proteinase activities in the midgut of Cry1Ab susceptible ECB larvae were carried out. Besides, the proteinases were tested for involvement in the digestion of the B.t.-toxin Cry1Ab and the respective protoxin.
Materials and methods
Isolation of midgut sap and BBMV ECB larvae were reared on artificial diet up to the 5th instar. For the extraction of both, the pure midgut sap and the midgut epithelium, the larvae were calmed on ice and dissected. The total midguts were isolated and collected on ice. Due to the very small sizes of 1st and 2nd instar larvae it was not possible to separate their midguts. For the sap extraction the midguts as well as crushed whole 1st and 2nd instar larvae were centrifuged at 13.000 g for 15 min. The resuite was stored at -18°C until usage. For preparation of brush border membrane vesicles (BBMV) the isolated midguts were treated as described by Wolfersberger et al. (1987). Photometrical tests For the identification and quantification of proteinases, photometrical studies were conducted using typical proteinase-indicating chromogenic substrates and specific inhibitors according to the investigations of Wagner et al. (2002): Trypsin was tested with the substrate N- benzoyl-L-arg-p-nitroanilide (BApNA) and soybean-trypsin-inhibitor (SBTI). Chymotrypsin was tested with N-succinyl-ala-ala-phe-p-nitroanilide (SAAFpNA) and the inhibitor N-tosyl- L-phe chloromethylketone (TPCK). Elastase was tested with N-succinyl-ala-ala-pro-leu-p- nitroanilide (SAAPLpNA) and elastatinal. Aminopeptidase was tested with leu-p-nitroanilide (LpNA) and bestatin. Carboxypeptidase was tested with hippuryl-phe and hippuryl-arg. Proteolytic assays and SDS-PAGE The experiments were done with purified Cry1Ab toxin and protoxin which was prepared by Dr. J.A. Jehle (State Education and Research Center for Agriculture, Viticulture and Horticulture; SLFA Neustadt; Germany). Model proteinases were obtained from Sigma. Digestions were performed at 25°C with a final toxin concentration of 1 mg/ml and either a midgut sap dilution of 1:10 or a model proteinase concentration of 0.25 mg/ml, respectively. Proteolyses were stopped by heating the samples for 2 min at 95°C.
SDS-PAGE was done according to Laemmli (1970) using the Roti-Load1 no. K929.1
denaturation buffer from Roth and 15% polyacrylamide gels. Fluka standards no. 69810 (indicated as "low") and 69811 (indicated as "high") were used as reference standards. Results and discussion
Proteinase activity in the midgut sap of 5th instar larvae In the midgut of a Canadian ECB population, Houseman & Chin (1995) identified the digestive proteinases trypsin, chymotrypsin, elastase, and aminopeptidase. To compare their results with German ECB, a reference system, which is also intended to be used to characterize potential available resistant ECB's, was established to identify and to quantify changes in proteinase-activities in the midgut of the pest insect. Thus, midgut sap of German susceptible 5th instar larvae was extracted and photometrical studies were carried out. Similar to the above described results, trypsin, chymotrypsin, elastase, and aminopeptidase were identified (Kaiser-Alexnat et al., 2003). In additional tests the presence of other potential activities, e.g. carboxypeptidase, could not be highlighted (data not shown).
Beside the examined serine proteinases and metalloproteinases, other classes of pro-
teolytic activities are unlikely to be present in the midgut sap of ECB due to physiological reasons. As reviewed by Terra et al. (1996), cysteine proteinases are generally common in the midgut of hemipteran Heteroptera or in slightly acid media and aspartic proteinases are only active at very acid pH values. Kaiser-Alexnat et al. (2003) demonstrated that the pH of pure
larval midgut sap of ECB 5th instar larvae is lightly basic, ranging between 7.2 and 7.5, depending on the rearing method before sample preparation. Proteinase activityin whole 1st and 2nd instar larvae Generally, early larval stages are known to be more sensitive to B.t.-toxin than late instars. Unfortunately, it was not possible to separate the midguts of 1st and 2nd instar larvae due to their very small sizes. Thus, with sap of whole larvae it was examined whether the activity of the above described proteinases is provable, too. As a control, no proteinase activity could be demonstrated in the haemolymphe (data not shown). In the sap of whole 1st (Fig. 1) and 2nd (Fig. 2) instar larvae, aminopeptidase activity was identified and quantified in photometrical tests. The diagrams show the means and the standard deviation of each experiment which was done three times. Columns indicated as "0" quantify the prevailing proteolytic activity; columns indicated with an increasing concentration of inhibitor show the specific inhibition of the proteolyses which is a tool to identify the type of proteinase.
Figure 1. Aminopeptidase-activity (± sd) in the sap of whole 1st instar larvae.
Figure 2. Aminopeptidase-activity (± sd) in the sap of whole 2nd instar larvae. Proteinase activityof BBMV from 5th instar larvae A membrane-bound aminopeptidase is one possible receptor for B.t.-toxins in the midgut epithelium (Oppert, 1999). Based on this fact and in context with the above described results,
aminopeptidase activity of BBMV isolated from 5th instar larvae was demonstrated using the established test system (Fig. 3). Binding analyses are presently carried out to show the inter-action between Cry1Ab and the receptor.
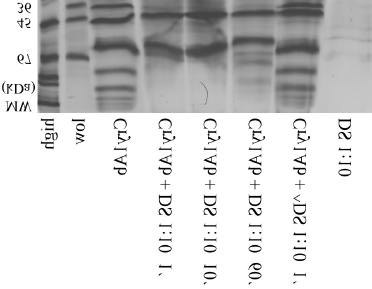
Figure 3. Aminopeptidase-activity (± sd) in the midgut epithelium (BBMV's) of 5th instar larvae. Proteolytical processing of B.t. toxin and protoxin Cry1Ab After incubation with midgut sap from 5th instar larvae, the B.t.-toxin Cry1Ab was processed during the first minute. As a result, the 65 kDa toxin was digested for 2 kDa, resulting in a 63 kDa protein. This protein is stable for at least 60 minutes (Fig. 4). In order to show that this protein shortening is due to proteolytic activity, a control was performed using midgut sap that was heated at 95°C for 5 min. Due to this denaturation of the proteinases, no digestion of the B.t.-toxin took place (see also Fig. 4).
Figure 4. Proteolytical processing of B.t.-toxin Cry1Ab with midgut sap (DS) of 5th instar larvae.
To identify the activity among the midgut proteinases, which are responsible for the pro-teolytical reaction, available model proteinases were used to simulate the midgut conditions, i.e. bovine trypsin, bovine chymotrypsin, porcine elastase, and Aeromonas aminopeptidase. As a result, both the 65 kDa toxin as well as the 135 kDa protoxin were digested to 63 kDa by all types of proteinases proved in the midgut sap of ECB, except aminopeptidase. The pro-teolytical processing with trypsin and chymotrypsin is demonstrated in Fig. 5 and the one with elastase and aminopeptidase in Fig. 6. Figure 5. Proteolytical processing with trypsin and chymotrypsin.
Figure 6. Proteolytical processing with elastase and aminopeptidase.
Acknowledgements
Thanks are due to Dr. J.A. Jehle, State Education and Research Center for Agriculture, Viticulture and Horticulture (SLFA Neustadt), Biotechnological Crop Protection, Neustadt/Weinstrasse, Germany for providing us with Cry1Ab toxin and protoxin. Thanks are also due to the BMBF for supporting the research project. References
Ferré, J. & van Rie, J. 2002: Biochemistry and genetics of insect resistance to Bacillus thuringiensis. Annu. Rev. Entomol. 47: 501-533.
Houseman, J.G. & Chin, P.-S. 1995: Distribution of digestive proteinases in the alimentary
tract of the European corn borer Ostrinia nubilalis (Lepidoptera: Pyralidae). Arch. Insect Biochem. Physiol. 28: 103-111.
Kaiser-Alexnat, R., Wagner, W., Langenbruch, G.A., Kleespies, R.G., Keller, B., Meise, T.,
& Hommel, B. 2003: Selection of resistant European corn borer (Ostrinia nubilalis) to B.t.-corn and preliminary studies for the biochemical characterization. IOBC wprs Bulletin. 9th European Meeting of the IOBC wprs Working goup "Insect pathogens and entomoparasitic nematodes" Growing biocontrol markets challenge research and development. May 23-29, 2003, Schloß Salzau, Germany, in press.
Laemmli, U.K. 1970: Cleavage of structural proteins during the assembly of the head of
McGaughey, W.H. & Oppert, B. 1998: Mechanisms of insect resistance to Bacillus thuringiensis toxins. Israel J. Entomol. 32: 1-14.
Oppert, B., Kramer, K.J., & McGaughey, W.H. 1997: Insect resistance to Bacillus thuringien-
Oppert, B. 1999: Review: Protease interactions with Bacillus thuringiensis insecticidal toxins.
Arch. Insect Biochem. Physiol. 42: 1-12.
Terra, W.R., Ferreira, C., Joradâo, B.P., & Dillon, R.J. 1996: Digestive enzymes. In: Biology
of the insect midgut, eds. Lehane and Billingsley. Chapman and Hall, London: 153-166.
Wagner, W., Möhrlen, F., & Schnetter, W. 2002: Characterization of the proteolytic enzymes
in the midgut of the European Cockchafer, Melolontha melolontha (Coleoptera: Scarabaeidae). Insect Biochem. Molec. Biol. 32: 803-814.
Wolfersberger, M.G., Luthy, P., Parenti, P., Sacchi, V.F., Giordana, B., & Hanozet, G.M.
1987: Preparation and partial characterization of amino acid transporting brush border membrane vesicles from the larval midgut of the cabbage butterfly (Pieris brassicae). Comp. Biochem. Physiol. 86A: 301-308.
INSECTOX CRAWLING INSECT KILLER 1. IDENTIFICATION OF THE SUBSTANCE /PREPARATION AND OF THE COMPANY /UNDERTAKING and 11/03/200-24692 (fundamation and method of MSDS : F+ Extremely Flammable. Keep away from sources of ignition.Do not store at temperatures above 50 C. Do not smoke when using. HEALTH HAZARDS Xn Harmful may cause lung damage if swallowed. Keep out of reach of :N Very toxic to
Antibiotic Formulary Prescribing Advice Adult V6.4 Printed copies are not controlled and are valid on date of printing only. This version was last printed: 16 Apr. 12 Lower Respiratory Tract Infections Inc. COPD, Pneumonia, TB 4.3.1 Bronchitis, Acute 4.3.2 Bronchitis, Chronic And COPD, Acute Exacerbations Of Doxycycline 200mg loading dose then 100mg od po Amoxicillin 500mg po
 aminopeptidase activity of BBMV isolated from 5th instar larvae was demonstrated using the established test system (Fig. 3). Binding analyses are presently carried out to show the inter-action between Cry1Ab and the receptor.
aminopeptidase activity of BBMV isolated from 5th instar larvae was demonstrated using the established test system (Fig. 3). Binding analyses are presently carried out to show the inter-action between Cry1Ab and the receptor.

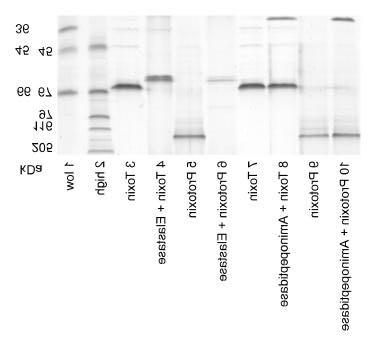 To identify the activity among the midgut proteinases, which are responsible for the pro-teolytical reaction, available model proteinases were used to simulate the midgut conditions, i.e. bovine trypsin, bovine chymotrypsin, porcine elastase, and Aeromonas aminopeptidase. As a result, both the 65 kDa toxin as well as the 135 kDa protoxin were digested to 63 kDa by all types of proteinases proved in the midgut sap of ECB, except aminopeptidase. The pro-teolytical processing with trypsin and chymotrypsin is demonstrated in Fig. 5 and the one with elastase and aminopeptidase in Fig. 6. Figure 5. Proteolytical processing with trypsin and chymotrypsin.
Figure 6. Proteolytical processing with elastase and aminopeptidase.
Acknowledgements
To identify the activity among the midgut proteinases, which are responsible for the pro-teolytical reaction, available model proteinases were used to simulate the midgut conditions, i.e. bovine trypsin, bovine chymotrypsin, porcine elastase, and Aeromonas aminopeptidase. As a result, both the 65 kDa toxin as well as the 135 kDa protoxin were digested to 63 kDa by all types of proteinases proved in the midgut sap of ECB, except aminopeptidase. The pro-teolytical processing with trypsin and chymotrypsin is demonstrated in Fig. 5 and the one with elastase and aminopeptidase in Fig. 6. Figure 5. Proteolytical processing with trypsin and chymotrypsin.
Figure 6. Proteolytical processing with elastase and aminopeptidase.
Acknowledgements