L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
Pii: s0006-2952(02)01121-8
Biochemical Pharmacology 64 (2002) 355±361
Department of Molecular and Cellular Biology, Ligand Pharmaceuticals, 10275 Science Center Drive, San Diego, CA 92121, USA
Glucocorticoids are the most effective anti-in¯ammatory agents known. However,the use of these powerful molecules is plagued by a
host of serious,sometimes life-threatening side-effects. The search for new compounds that maintain the ef®cacy of the steroids without
some of the side-effects has entered a new phase. New approaches are leading to novel kinds of steroidal and non-steroidal compounds
with unique pro®les that may represent the next generation of safer glucocorticoids.
# 2002 Elsevier Science Inc. All rights reserved.
Keywords: Selective glucocorticoid receptor modulators; Steroid receptor ligand; Non-steroidal anti-in¯ammatory agent
nately,early on,it was discovered that these compounds
had a severely negative impact on patients to whom they
Early efforts to understand endocrine function focused
were administered. Efforts from that point on have focused
on extracts derived from glands whose removal caused
on ®nding molecules that have anti-in¯ammatory ef®cacy
speci®c systemic pathologies in animals. The adrenal was
equal to that of the steroids,but with a reduction in side-
one such gland. Organic extracts from this gland could be
effects. Progress has been made with several synthetic
used to ameliorate the symptoms of Addison's disease
steroidal versions that exhibit increased receptor speci®-
(later discovered to be caused by the speci®c lack of the
city and potency (dexamethasone) as well as versions for
glucocorticoid hormone cortisol). Addison's patients exhi-
use in topical,inhaled,or other non-oral formulations.
bit hyperpigmentation of the skin,hypoglycemia,and salt
These,typically,are extremely potent steroids with high
craving,among other symptoms. These patients do not
ef®cacy that are cleared rapidly by ®rst-pass metabolism.
respond effectively to stress,creating the potential for an
This pro®le results in excellent ef®cacy at the point of
Addisonian crisis,a disease cascade that is potentially life-
application (e.g. lung and skin) but with reduced,although
threatening. The majority of these symptoms can be elimi-
nated with the injection of adrenal extracts.
The isolation of the active components from this gland
by Tadeus Reichstein and Edward Kendall,and the sub-
sequent use in patients with rheumatoid arthritis by Philip
Hench,garnered all three the Nobel Prize in 1950. These
Corticosteroids produced in the adrenal gland undergo
agents were remarkably effective at inhibiting many forms
metabolism into two compounds (glucocorticoids and
of in¯ammation and were used at high doses over long
mineralocorticoids) with markedly different activities;
periods of time,resulting in excellent ef®cacy. Unfortu-
glucocorticoids like cortisol have effects on carbohydrate,
fat,and protein metabolism,and mineralocorticoids like
aldosterone have effects on sodium levels by raising
Tel.: 1-858-550-7807; fax: 1-858-550-7235.
reabsorption in the kidney The responses to these
E-mail address: jminer@ligand.com (J.N. Miner).
Abbreviations: CRH,corticotropin-releasing hormone; CRF,corticotro-
hormones are mediated by two different intracellular
pin-releasing factor; ACTH,adrenocorticotropin hormone; HPA,hypotha-
receptors,which are capable of binding to and thereby
lamic-pituitary-adrenal; GVHD,graft-versus-host disease; PEPCK,
being activated by these steroids,translocating to the
phosphoenol pyruvate carboxy kinase; LBD,ligand-binding domain;
nucleus,and regulating speci®c target genes. This signal
GRIP-1,GR interacting protein 1; NFkB,nuclear factor-kappa B; AP-1,
activator protein 1; PPAR,peroxisome proliferator activated receptor; PGC-
transduction pathway is unique in that it utilizes a single
1,PPARg coactivator 1; HNF-4,hepatocyte nuclear factor 4.
protein that is both the proximal ligand receptor as well as
0006-2952/02/$ ± see front matter # 2002 Elsevier Science Inc. All rights reserved.
PII: S 0 0 0 6 - 2 9 5 2 ( 0 2 ) 0 1 1 2 1 - 8
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
the transcription factor that interacts with DNA in the
patients present with a combination of symptoms grouped
nucleus. The GR,in particular,is responsible for up-
under the heading `Cushing's syndrome' The symp-
regulating and down-regulating a wide range of genes
toms include central obesity,glucose intolerance,myopa-
affecting a number of critical metabolic pathways. Glu-
thy,and hypertension,among others. The pathophysiology
cocorticoids are commonly known as the stress hormones,
of this condition is most often related to dysregulation of
and,under normal circumstances,they are crucial to the
the system that produces glucocorticoids due to tumors or
ability of the body to respond and adapt to stress. Stress
other endocrine problems. Certain adrenal tumors are
causes signaling within the two key components of the
known to secrete large quantities of cortisol and are not
stress response,CRH neurons found in the paraventricular
regulated by normal feedback mechanisms. Over-secretion
nucleus of the hypothalamus and in other areas of the
of signaling molecules within the glucocorticoid produc-
central nervous system as well as the noradrenergic neu-
tion pathway,CRF or ACTH by the pituitary or non-
rons of the locus-ceruleus-norepinephrine system. Activa-
pituitary cells,can also increase serum levels of cortisol
tion of these systems induces psychological,behavioral,
and physical changes that provide an adaptive bene®t in the
The last and perhaps the most common way in which
short term. The HPA axis is on the receiving end of signals
imbalances in the stress response system can present
from the CRH neurons. This axis is the primary regulator
themselves is when patients are administered exogenous
of endogenous glucocorticoid production. CRF from CRH
neurons acts on the anterior pituitary gland to cause the
Glucocorticoids are extremely effective and frequently
secretion of ACTH,which,in turn,induces corticosteroid
used therapeutic agents that are administered for a wide
production and release from the adrenal gland. Thus,in
range of disorders. Supplemental glucocorticoids can
response to stress,the HPA axis is activated,and gluco-
replace cortisol absent in various adrenal insuf®ciency
corticoid levels in serum increase. The behavioral and
syndromes These compounds are also effective
psychological changes that occur when cortisol levels rise
anti-in¯ammatory agents for many autoimmune and
are indicative of a ®ght or ¯ight response and include
in¯ammatory disorders,such as rheumatoid arthritis and
increases in alertness,mental acuity,pain tolerance,tem-
asthma. With rheumatoid arthritis,treatment with non-
perature,and euphoria coupled with decreases in sexual
steroidal anti-in¯ammatory drugs (NSAIDs) provides sig-
desire,reproductive behavior,and appetite The phy-
ni®cant bene®t; however,the progression of the disease
sical/physiological changes include increases in respiration
eventually demands the use of glucocorticoids. Steroids
rate,oxygenation,cardiovascular tone,blood ¯ow,pres-
inhibit the signs and symptoms of the in¯ammation asso-
sure,and heart rate. Accompanying these are adjustments
ciated with rheumatoid arthritis,but fail to reverse any
in metabolism to provide glucose and energy by increasing
structural damage that has already occurred in the joint.
gluconeogenesis in the liver and lipolysis in fat depots.
Additionally,patients with asthma often use inhaled and
Protein is broken down to provide substrates for gluconeo-
oral steroids to control exacerbation of their condition.
genesis. A number of peripheral systems unnecessary to the
Immunosuppressive therapy for transplant rejection and
short-term stress response are inhibited. These suppressed
autoimmune disorders often makes use of short-term,high-
functions include growth,reproduction,food absorption,
dose treatment with steroids followed by more protracted
and the immune response. Interestingly,the stress response
lower dose treatment to reduce the cell-mediated response
also provides an autoregulatory mechanism to reduce the
to transplanted foreign tissue. Patients receiving bone
production of glucocorticoid after high levels have been
marrow transplants occasionally develop acute GVHD.
achieved. This is accomplished by feedback of glucocorti-
Glucocorticoids are extremely useful in GVHD; other
coids onto the signaling pathways that induce glucocorti-
therapeutics are used only for steroid-resistant GVHD
coid production. Glucocorticoids inhibit the expression of
genes involved in activating the HPA axis,including
multiple myeloma and certain lymphomas and leukemias,
corticotropin-releasing factor and ACTH,thereby regulat-
respond well to combination therapies that include the
ing corticosteroid production from the adrenal gland
glucocorticoids prednisone or dexamethasone.
There are several situations when this carefully balanced
Exposure to high,sustained levels of corticosteroids by
system can go awry. Chronic,uncontrolled stress leads to
any mechanism uncouples the normal metabolic processes
long-term activation of the HPA axis and sustained,high
from autoregulatory feedback mechanisms and induces a
glucocorticoid levels. Pathological conditions in which
stress response physiological state that cannot be main-
activation of the HPA axis has been demonstrated include
tained long term without severe consequences. The numer-
depression,obsessiveÐcompulsive disorder,alcohol and
ous side-effects experienced by patients administered
drug abuse,and anorexia nervosa The action of
steroids over the long term are perhaps the clearest exam-
glucocorticoids in response to chronic stress is decidedly
ple of this. The response to glucocorticoids is complex,due
in large measure to the wide variety of physiological
Imbalances in cortisol production can also occur in
contexts in which glucocorticoids act. Complications
certain conditions that overproduce glucocorticoids. These
are time- and dose-dependent and can occur acutely with
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
very high doses,or more slowly with chronic exposure and
the glucose production pathways. While bene®cial in the
lower doses. Fortunately,glucocorticoid-regulated clinical
short term,this decreases overall muscle function when
markers of speci®c side-effects are available from the
activated for an extended period of time.
extensive clinical trials conducted over the years with
The behavioral effects of glucocorticoids are of great
glucocorticoids. These markers are relevant to many of
concern to patients. Glucocorticoids have long been known
the impacts of glucocorticoids on bone,fat,and carbohy-
to have psychogenic effects in a subset of patients when
drate metabolism. Most are readily monitored in response
given at high doses. Approximately 5% of patients will
to short-term exposure to glucocorticoids and include
experience some degree of inappropriate euphoria,psy-
serum cortisol,a measure of HPA suppression,lipid pro®le
chosis,or depression Patients are also quite con-
changes,which measure weight effects,serum glucose and
cerned with the effects of glucocorticoids on fat
insulin levels,a measure of effects on insulin resistance
redistribution and weight gain. Fat and muscle are lost
and hepatic glucose output,and urinary collagen peptides
from limbs,but truncal and visceral areas actually accu-
and serum osteocalcin that help assess the impact of
mulate fat. Facial,supraclavical,and posterior cervical fat
compounds on bone metabolism. Together,these markers
depots are particularly sensitive to glucocorticoids,result-
can assist in the assessment of speci®c compounds early in
ing in the moon face and buffalo hump characteristic of
clinical trials. The speci®c side-effects of glucocorticoids
long-term glucocorticoid treatment Even one dose of
can be ranked by patients and physicians. These vary
a glucocorticoid is suf®cient to increase hepatic glucose
somewhat depending on the person doing the ranking.
production and increase insulin resistance of peripheral
In particular,patients tend to highlight the physical and
tissues. The glucocorticoid effect on glycemic control is
mental changes that accompany long-term steroid use.
thought to target insulin signaling. Glucocorticoids
These include fat redistribution and weight gain and
affect insulin-mediated increases in blood ¯ow to muscles
steroid-induced psychosis/neurosis. Physicians,on the
They decrease key insulin receptor signaling mole-
other hand,tend to concentrate on problems that affect
cules and increase glucose output by increasing the rate-
patient medical care,which include hyperglycemia,gen-
limiting enzyme in gluconeogenesis,PEPCK.
eralized insulin resistance,as well as suppression of the
Glucocorticoids also inhibit the release of insulin from
HPA axis. However,without question,the single most
the pancreas,acting directly on the pancreatic b cells. This
important side-effect from the standpoint of many physi-
action may involve apoptosis of b cell populations,leading
cians is osteoporosis. This side-effect alone accounts for an
to decreased insulin production as well as more direct
enormous amount of morbidity among patients receiving
inhibition of insulin expression The molecular
glucocorticoids. Long-term glucocorticoid treatment often
details underpinning regulation of hepatic glucose produc-
results in some degree of osteoporosis in patients. Because
tion have been made clearer recently by discoveries linking
these patients also suffer from decreased muscle mass as a
GR,other transcription factors,and cyclic AMP (cAMP) in
consequence of steroid treatment,they are more suscep-
tible to falling. The consequences of falls and subsequent
fractures become enormous when one considers that many
of these patients are already sick with a debilitating dis-
ease. A hip fracture late in life with its attendant inactivity
and increased potential for pneumonia can have fatal
The GR acts as a ligand-regulated transcription factor
consequences for elderly patients. Susceptibility to frac-
responding to circulating cortisol. It is a member of the
tures and aseptic necrosis of the femoral head increases
large family of intracellular receptors comprised of both
within months of starting glucocorticoid therapy
the nuclear hormone receptors as well as the steroid
Steroids degrade the quality of trabecular bone,resulting in
receptor subfamilies. The protein itself is composed of
an increase in fracture rate. Bone loss is highest in the
three general domains: a DNA-binding domain,a C-term-
®rst 6 months of therapy,after which patients continue to
inal ligand-binding domain (LBD),and an N-terminal
lose bone,but at a slower rate. When taken off steroids,
activation domain. It is capable of regulating transcription
patients do appear to partially regain bone The loss
both negatively and positively and is localized to the
of muscle compounds the osteoporosis problem. Gluco-
cytoplasm in the absence of hormone. The receptor is held
corticoid-induced myopathy results in decreased strength
in an inactive state poised to bind ligand by interaction with
and muscle mass. The mechanism by which glucocorti-
a chaperonin complex comprised of heat shock proteins.
coids affect muscle mass is partially due to hypogonadism
Upon binding ligand,the receptor undergoes a conforma-
observed in many patients with the consequent decline in
tional change that dissociates the heat shock proteins and
levels of the sex steroids estrogen and testosterone,which
activates a number of receptor functions including DNA
are responsible for contributing to the maintenance of both
binding activity,nuclear localization,and transcriptional
muscle and bone mass Furthermore,mimicking a
regulation. The latter involves the direct and indirect inter-
stress response,muscle is broken down and utilized as a
action with a large number of transcription factors critical
source of substrates and energy for the increased activity in
to gene regulation including RNA polymerase as well
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
as various polymerase-associated proteins. The complex
repression prompted a recent analysis of the collage-
that forms at a regulated gene is quite large,and the
nase-3 gene promoter under glucocorticoid-repressed con-
receptor likely does not contact RNA polymerase directly,
ditions Given the previously described role for GRIP-1
but instead utilizes several types of so-called ``coactivator''
as a coactivator,it came as a surprise when these authors
proteins to bridge the gap between itself and the polymer-
demonstrated an important role for GRIP-1 in repression.
ase The details of the interaction between the receptor
They demonstrated that GRIP was recruited to the col-
and these coactivators are understood from genetic,bio-
lagenase promoter during the act of repression by the GR.
chemical,and crystallographic standpoints for only a few
The ligand-dependent recruitment of these proteins might
speci®c receptorÐcoactivator pairs. In general,these inter-
be used as a readout to detect potentially bene®cial ther-
actions make use of the LBD of the receptor,although
other less well-characterized interactions clearly occur in
The search for a novel glucocorticoid that has the anti-
the N-terminal and DNA binding domains Coactiva-
in¯ammatory properties of conventional steroids without
tors bind to the LBD of the GR in a hormone-dependent
one or more of the side-effects has been a long-standing
fashion,interacting directly with the extreme C-terminal
goal of the ®eld. Much effort has been spent on modifying
portion of the LBD. This interaction domain is formed by
the steroid backbone to achieve this sort of increased
the juxtaposition of several helical segments within the
therapeutic index; however,these efforts have met with
protein. The interaction surface between these transcrip-
little success. De¯azacort,a D-ring-substituted steroid
tion factors is comprised of a hydrophobic pocket on the
otherwise similar to cortisol,was touted originally as a
receptor and a helical sequence containing an LxxLL
powerful anti-in¯ammatory molecule exhibiting more
amino acid motif on the coactivator This interaction
selective,i.e. reduced,activity,in particular on bone and
is very sensitive to the structure of the ligand bound in the
on glucose metabolism. Initially,clinical data supported
pocket. Coactivators typically bind avidly in the presence
this notion. However,subsequent trials that adjusted
of agonists,but fail to bind in the presence of antagonist
the steroid dose to maintain equivalent anti-in¯ammatory
ligands. In fact,this is likely the mechanism of antagonist
ef®cacy usually needed higher levels of de¯azacort. Unfor-
action. This conformational sensitivity is due to the
tunately,at these higher doses,the advantages of de¯azacort
changes in receptor structure brought about by the structure
disappeared The ®eld was re-energized by the dis-
of the ligand bound in the pocket. The receptor appears to
covery of the likely mechanism of GR-mediated repression
actually condense around the ligand during the binding
of a wide variety of pro-in¯ammatory genes. The receptor
reaction,meaning that structural changes in the ligand are
was shown to bind directly to speci®c transcription factors
transmitted directly to the receptor and to the receptor
(NFkB and AP-1) involved in up-regulating in¯ammatory
surfaces that interact with coactivators Thus,ligands
genes. This represented a unique mechanism that was
can change the ability of the receptor to bind to coactivators.
genetically separable from transcriptional activation. The
These coactivators may also play a role in the tissue-speci®c
search began for ligands that could induce transcriptional
activity of glucocorticoids. Although many coactivators are
repression,but hinder transcriptional activation. In 1997,
expressed widely,some have been described that exhibit a
the ®rst compounds that separated transactivation from
restricted tissue expression pattern Coactivators are
not only involved in transcriptional activation,but they also
These compounds were steroidal in nature,were very
appear to play a role in transcriptional repression. Recent
ef®cient inhibitors of both AP-1- and NFkB-mediated gene
data have demonstrated that transcriptional repression by
induction,and were strong anti-in¯ammatory agents in
many members of the nuclear hormone receptor family is
vivo. They also were reported to have reduced ability to
dependent upon speci®c corepressor proteins that bind
activate gene expression in some,but not all cellular
directly to the receptor in the absence of hormone and
contexts. Unfortunately,thus far,no in vivo therapeutic
inhibit the transcription process by recruiting histone dea-
advantage has been demonstrated for these types of mole-
cetylases ,which appear to condense chromatin and
cules when side-effects were measured This result
therefore repress transcription. However,most mem-
calls into question the usefulness of the activation±repres-
bers of the steroid receptor subfamily are sequestered in the
cytoplasm in the absence of hormone and exhibit gene-
We have also pursued this hypothesis as an approach to
speci®c repression or activation only when hormone is
discovering selective GR modulators. An example of the
present. Corepressors have not yet been shown to bind
compounds identi®ed during this collaborative effort
steroid receptors during normal gene regulation
between Ligand Pharmaceuticals and Abbott Laboratories
although there is evidence for interaction when bound to
is AL-438 (Abbott-Ligand 438),which was shown to be a
speci®c,non-steroidal ligand for the GR that exhibited a
The ubiquitous coactivator GRIP-1 has been shown to
unique pro®le,both in vitro and in vivo. The molecule is
bind to GR and other intracellular receptors and to enhance
fully ef®cacious at transcriptional repression compared
their transcriptional activation activity. Unresolved
with prednisolone on certain genes related to the anti-
questions regarding the mechanism of steroid receptor
in¯ammatory aspects of glucocorticoid activity (E-selectin
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
and interleukin-6),but is weaker (a partial agonist) for
This is consistent with the gene-speci®c activity
transcriptional activation. Using other promoters in differ-
described earlier. The mechanism may be found in the
ent cell backgrounds,AL-438 is more active as a transcrip-
fact that AL-438 generates a receptor conformation that
tional activator. Thus,AL-438 does not completely
differs from steroids,which in turn changes the spectrum of
separate transcriptional repression from activation,but
coactivators with which the receptor can interact. Certain
instead appears to be separating activities in a gene-speci®c
coactivators bind GR with identical af®nity in the presence
fashion. In animal models,AL-438 was as ef®cacious as
of either AL-438 or prednisolone,while others exhibit
prednisolone at inhibiting in¯ammation. Importantly,AL-
signi®cantly reduced af®nity in the presence of this com-
438 exhibited signi®cantly reduced impact on fasting
pound. We believe that since different genes have different
glucose levels compared with prednisolone,suggesting
requirements for speci®c coactivators,this may be the
that this compound might not cause the diabetogenic
molecular rationale for AL-438's gene-selective pro®le.
effects of steroidal glucocorticoids.
The coactivator GRIP-1 is an example of a potential
An as-yet-unanswered question is whether the improved
mediator of gene-speci®c effects,given its demonstrated
pro®le of AL-438 in vivo is a direct result of its altered effect
involvement in both transcriptional repression and activa-
on GR structure and function as detected by in vitro assays.
tion. We have examined the interactions between GRIP-1
It is possible that it is the speci®c conformation of the
and GR in a variety of mammalian two-hybrid and GST
receptor detected by our in vitro assays that is responsible
pull-down assays. Our ®ndings indicate that,in a manner
for the therapeutically bene®cial pro®le observed in rodents.
similar to prednisolone,AL-438 will induce the interaction
Fig. 1. Selective glucocorticoid receptor modulator (SGRM) coactivator hypothesis. This figure depicts a model that attempts to distinguish the effects of
steroidal glucocorticoids from SGRMs. Both compounds are fully efficacious at binding GR and causing the interaction with GRIP-1,a coactivator involved
in repression of inflammatory genes by GR. The model suggests that this interaction is,in part,responsible for the anti-inflammatory activity of both
compounds. Of course this is likely not the entire explanation. That said,when examining the ability of steroids and SGRMs to induce the interaction with
PGC-1,a coactivator involved in glucose homeostasis in the liver among other things,a strikingly different result is found. The SGRM is weaker at inducing
the PGC-1 interaction than the steroid. This may help explain the reduced negative impact on glucose metabolism from the SGRM. Thus,the reduced PGC-1
interaction may translate into reduced side-effects in liver and possibly other tissues as well.
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
between GRIP and GR ef®ciently. This ®nding suggests
exhibitactivitiesthathavethepromiseofleadingtopowerful,
that the differences between AL-438 and steroids in vivo
yet safer GR-dependent anti-in¯ammatory agents.
are not to be explained by the GRIP-1ÐGR interaction,
with the caveat that we have not tested all cell contexts with
this approach. Another example of potential coactivators
for the GR is the PGC-1,originally characterized as a
PPAR coactivator highly expressed in brown fat,involved
[1] Rohatagi S,Rhodes GR,Chaikin P. Absolute oral versus inhaled
in fat differentiation ,and more recently shown to play
bioavailability: significance for inhaled drugs with special reference to
inhaled glucocorticoids. J Clin Pharmacol 1999;39:661±3.
a critical role in glucocorticoid-mediated stimulation of
[2] Mackie AE,Bye A. The relationship between systemic exposure to
glucose production from the liver. The PEPCK gene
fluticasone propionate and cortisol reduction in healthy male volun-
requires the action of PGC-1 together with the transcription
teers. Clin Pharmacokinet 2000;39:47±54.
factor HNF-4 to ef®ciently respond to glucocorticoids
[3] Orth DN,Kovacs WJ. The adrenal cortex. In: Wilson JD,Foster DW,
These authors demonstrate that PGC-1 binds
Kronenberg HM,Larsen PR,editors. Williams textbook of endocri-
nology. Philadelphia: Saunders,1998. p. 517.
directly to GR and to HNF-4 and is critical for the appro-
[4] Peeke PM,Chrousos GP. Hypercortisolism and obesity. Ann NYAcad
priate response to both cAMP signaling and glucocorti-
coids. We became interested in this protein initially
[5] Chrousos GP. The role of stress and the hypothalamic-pituitary-
because of the critical role played by PGC-1 in fat meta-
adrenal axis in the pathogenesis of the metabolic syndrome: neuro-
bolism and differentiation activity intimately tied
endocrine and target tissue-related causes. Int J Obes Relat Metab
to the effects of glucocorticoids. The PGC-1ÐGR con-
[6] Gold PW,Gwirtsman H,Avgerinos PC,Nieman LK,Gallucci WT,
nection was strengthened considerably by the demonstra-
Kaye W,Jimerson D,Ebert M,Rittmaster R,Loriaux DL. Abnormal
tion of direct binding to GR and most recently by its
hypothalamic-pituitary-adrenal function in anorexia nervosa. Patho-
involvement in glucocorticoid effects on glucose produc-
physiologic mechanisms in underweight and weight-corrected pa-
tion. Thus,we hypothesized that the reduced
tients. N Engl J Med 1986;314:1335±42.
[7] Gold PW,Goodwin FK,Chrousos GP. Clinical and biochemical
impact of AL-438 on fat metabolism and glucose levels
manifestations of depression. Relation to the neurobiology of stress
compared with prednisolone might be connected to
(First of two parts). N Engl J Med 1988;319:348±53.
changes in the interaction with PGC-1. This notion was
[8] Norton JA,Li M,Gillary J,Le HN. Cushing's syndrome. Curr Probl
tested with direct interaction assays that demonstrated that
PGC-1 could bind directly to the GR in response to
[9] Boumpas DT,Chrousos GP,Wilder RL,Cupps TR,Balow JE.
Glucocorticoid therapy for immune-mediated diseases: basic and
prednisolone and dexamethasone. The response to AL-
clinical correlates. Ann Intern Med 1993;119:1198±208.
438 in the same assays was reduced,suggesting that,in
[10] Deeg HJ,Henslee-Downey PJ. Management of acute graft-versus-host
contrast to GRIP-1,AL-438 did not induce a GR con-
disease. Bone Marrow Transplant 1990;6:1±8.
formation that ef®ciently interacted with PGC-1.
[11] Schleimer RP,Freeland HS,Peters SP,Brown KE,Derse CP. An
outlines the hypothesis that these data supported.
assessment of the effects of glucocorticoids on degranulation,che-
This idea suggested that for the inhibition of in¯amma-
motaxis,binding to vascular endothelium and formation of leukotriene
tory mediators (collagenase,interleukin-6,and E-selectin),
4 by purified human neutrophils. J Pharmacol Exp Ther 1989;250:
interaction with GRIP-1 was important d both AL-
[12] Axelrod J,Reisine TD. Stress hormones: their interaction and regula-
438 and steroids induce this interaction ef®ciently. In
contrast,the interaction with PGC-1 is known to be
[13] Lane NE,Lukert B. The science and therapy of glucocorticoid-
important for the effects of glucocorticoids on liver glucose
induced bone loss. Endocrinol Metab Clin North Am 1998;27:465±83.
[14] Ralston SH. The genetics of osteoporosis. Bone 1999;25:85±6.
production and hyperglycemia AL-438 is less
[15] van Staa TP,Leufkens HGM,Abenhaim L,Zhang B,Cooper C. Oral
ef®cient at inducing PGC-1 interactions when compared
corticosteroids and fracture risk: relationship to daily and cumulative
with steroids,perhaps explaining the differences between
doses. Rheumatology (Oxford) 2000;39:1383±9.
the compound and the steroids on glucose levels in vivo.
[16] Crilly R,Cawood M,Marshall DH,Nordin BE. Hormonal status in
This hypothesis remains to be tested by directly demon-
normal,osteoporotic and corticosteroid-treated postmenopausal wo-
strating the involvement of PGC-1 and GRIP-1 in the
[17] Doerr P,Pirke KM. Cortisol-induced suppression of plasma testoster-
differential activity of AL-438. Furthermore,the differen-
one in normal adult males. J Clin Endocrinol Metab 1976;43:622±9.
tial interactions induced by these compounds may extend
[18] The Boston Collaborative Drug Surveillance Program. Acute adverse
reactions to prednisone in relation to dosage. Clin Pharmacol Ther
For the past 30 years,efforts to develop new,more power-
[19] Baxter JD. The effects of glucocorticoid therapy. Hosp Pract (Off Ed)
ful,and safer anti-in¯ammatory agents that work through the
GR have concentrated mainly on the steroid backbone.
[20] Tappy L,Randin D,Vollenweider P,Vollenweider L,Paquot N,
Recent discoveries of the molecular and cellular aspects of
Scherrer U,Schneiter P,Nicod P,Jequier E. Mechanisms of dexa-
GR activity have opened several exciting new approaches to
methasone-induced insulin resistance in healthy humans. J Clin En-
this effort. Indeed,recent advances in chemistry and screen-
[21] Laakso M,Edelman SV,Brechtel G,Baron AD. Decreased effect of
ing,together with new molecular approaches,have allowed
insulin to stimulate skeletal muscle blood flow in obese man. A novel
the identi®cation of novel non-steroidal compounds that
mechanism for insulin resistance. J Clin Invest 1990;85:1844±52.
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
[22] Sutherland C,O'Brien RM,Granner DK. New connections in the
[34] Wagner BL,Norris JD,Knotts TA,Weigel NL,McDonnell DP. The
regulation of PEPCK gene expression by insulin. Philos Trans R Soc
nuclear corepressors NCoR and SMRT are key regulators of both
ligand- and 8-bromo-cyclic AMP-dependent transcriptional activity of
[23] Granner DK,Sasaki K,Chu D. Multihormonal regulation of phos-
the human progesterone receptor. Mol Cell Biol 1998;18:1369±78.
phoenolpyruvate carboxykinase gene transcription. The dominant role
[35] Hong H,Kohli K,Trivedi A,Johnson DL,Stallcup MR. GRIP-1,a
of insulin. Ann NY Acad Sci 1986;478:175±90.
novel mouse protein that serves as a transcriptional coactivator in yeast
[24] Fernandez-Mejia C,Medina-Martinez O,Martinez-Perez L,Goodman
for the hormone binding domains of steroid receptors. Proc Natl Acad
PA. The human insulin gene contains multiple transcriptional ele-
ments that respond to glucocorticoids. Pancreas 1999;18:336±41.
[36] Rogatsky I,Zarember KA,Yamamoto KR. Factor recruitment and
[25] Delaunay F,Khan A,Cintra A,Davani B,Ling Z-C,Andersson A,
TIF2/GRIP-1 corepressor activity at a collagenase-3 response element
OÈstenson C-G,Gustafsson J-AÊ,Efendic S,Okret S. Pancreatic b cells
that mediates regulation by phorbol esters and hormones. EMBO J
are important targets for the diabetogenic effects of glucocorticoids. J
[37] Markham A,Bryson HM. Deflazacort. A review of its pharmacolo-
[26] Lambillotte C,Gilon P,Henquin J-C. Direct glucocorticoid inhibition
gical properties therapeutic efficacy. Drugs 1995;50:317±33.
of insulin secretion. An in vitro study of dexamethasone effects in
[38] Canalis E,Avioli L. Effects of deflazacort on aspects of bone forma-
mouse islets. J Clin Invest 1997;99:414±23.
tion in cultures of intact calvariae and osteoblast-enriched cells. J Bone
[27] Jenkins BD,Pullen CB,Darimont BD. Novel glucocorticoid receptor
coactivator effector mechanisms. Trends Endocrinol Metab
[39] VayssieÁre BM,Dupont S,Choquart A,Petit F,Garcia T,Marchandeau
C,Gronemeyer H,Resche-Rigon M. Synthetic glucocorticoids that
[28] Westin S,Rosenfeld MG,Glass CK. Nuclear receptor coactivators.
dissociate transactivation and AP-1 transrepression exhibit anti-in-
flammatory activity in vivo. Mol Endocrinol 1997;11:1245±55.
[29] Knutti D,Kralli A. PGC-1,a versatile coactivator. Trends Endocrinol
[40] Belvisi MG,Brown TJ,Wicks S,Foster ML. New glucocorticosteroids
with an improved therapeutic ratio? Pulm Pharmacol Ther 2001;14:
[30] Spiegelman BM,Puigserver P,Wu Z. Regulation of adipogenesis and
energy balance by PPARg and PGC-1. Int J Obes Relat Metab Disord
[41] Belvisi MG,Wicks SL,Battram CH,Bottoms SEW,Redford JE,
Woodman P,Brown TJ,Webber SE,Foster ML. Therapeutic benefit of
[31] Heinzel T,Lavinsky RM,Mullen T-M,SoÈderstroÈm M,Laherty CD,
a dissociated glucocorticoid and the relevance of in vitro separation of
Torchia J,Yang W-M,Brard G,Ngo SD,Davie JR,Seto E,Eisenman
RN,Rose DW,Glass CK,Rosenfeld MG. A complex containing N-
CoR,mSin3 and histone deacetylase mediates transcriptional repres-
[42] Herzig S,Long F,Jhala US,Hedrick S,Quinn R,Bauer A,Rudolph D,
Schutz G,Yoon C,Puigserver P,Spiegelman B,Montminy M. CREB
[32] Dressel U,Thormeyer D,Altincicek B,Paululat A,Eggert M,
regulates hepatic gluconeogenesis through the coactivator PGC-1.
Schneider S,Tenbaum SP,Renkawitz R,Baniahmad A. Alien,a
highly conserved protein with characteristics of a corepressor for
[43] Yoon JC,Puigserver P,Chen G,Donovan J,Wu Z,Rhee J,Adelmant
members of the nuclear hormone receptor superfamily. Mol Cell Biol
G,Stafford J,Kahn CR,Granner DK,Newgard CB,Spiegelman BM.
Control of hepatic gluconeogenesis through the transcriptional coac-
[33] Horlein AJ,Naar AM,Heinzel T,Torchia J,Gloss B,Kurokawa R,
tivator PGC-1. Nature 2001;413:131±8.
Ryan A,Kamei Y,Soderstrom M,Glass CK. Ligand-independent
[44] Knutti D,Kaul A,Kralli A. A tissue-specific coactivator of steroid
repression by the thyroid hormone receptor mediated by a nuclear
receptors,identified in a functional genetic screen. Mol Cell Biol
receptor co-repressor. Nature 1995;377:397±404.
Publikationsverzeichnis Prof. Dr. med. Anita Rauch A) Originalarbeiten 108. Göhring I, Tagariello A, Endele S, Stolt CC, Ghassibé M, Fisher M, Thiel CT, Trautmann U, Vikkula M, Winterpacht A, FitzPatrick DR, Rauch A (2009) Disruption of ST5 is associated with mental-retardation and multiple congenital anomalies. J Med Genet, in press 107. Segel R, Levy-Lahad E, Pasutto F, Picard
Newsletter Sections: Message from the President - Charles Bouloux Market Highlight - Marine / E-Marine The month of May was a time of focus and direction for our regional AIG Events in the Region Regional Management conference in Oman. The conference enforced our focus and direction on servicing our MEMSA Newsroom customer und

 Biochemical Pharmacology 64 (2002) 355±361
Department of Molecular and Cellular Biology, Ligand Pharmaceuticals, 10275 Science Center Drive, San Diego, CA 92121, USA
Glucocorticoids are the most effective anti-in¯ammatory agents known. However,the use of these powerful molecules is plagued by a
host of serious,sometimes life-threatening side-effects. The search for new compounds that maintain the ef®cacy of the steroids without
some of the side-effects has entered a new phase. New approaches are leading to novel kinds of steroidal and non-steroidal compounds
with unique pro®les that may represent the next generation of safer glucocorticoids.
Biochemical Pharmacology 64 (2002) 355±361
Department of Molecular and Cellular Biology, Ligand Pharmaceuticals, 10275 Science Center Drive, San Diego, CA 92121, USA
Glucocorticoids are the most effective anti-in¯ammatory agents known. However,the use of these powerful molecules is plagued by a
host of serious,sometimes life-threatening side-effects. The search for new compounds that maintain the ef®cacy of the steroids without
some of the side-effects has entered a new phase. New approaches are leading to novel kinds of steroidal and non-steroidal compounds
with unique pro®les that may represent the next generation of safer glucocorticoids.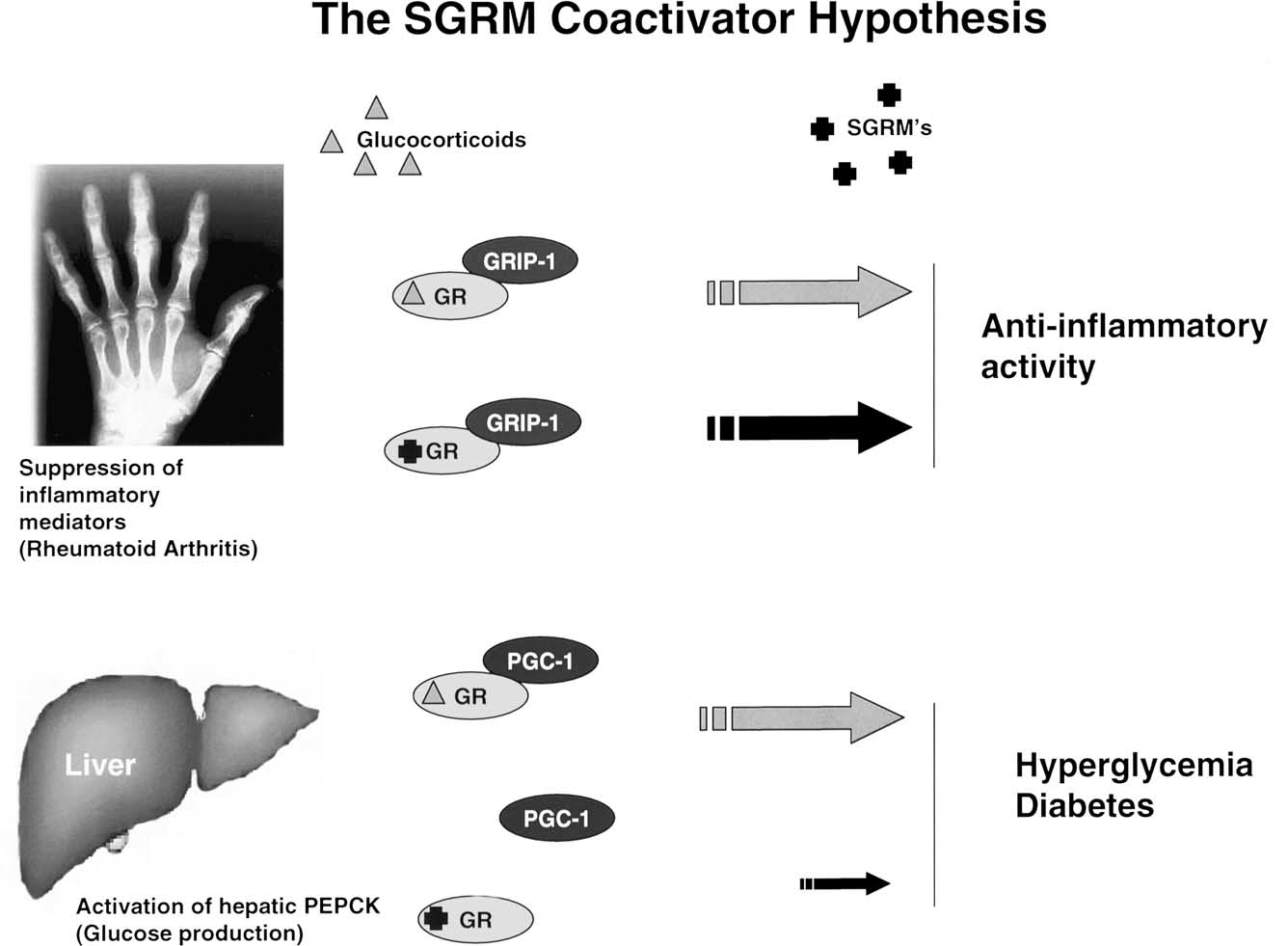 J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
and interleukin-6),but is weaker (a partial agonist) for
This is consistent with the gene-speci®c activity
transcriptional activation. Using other promoters in differ-
described earlier. The mechanism may be found in the
ent cell backgrounds,AL-438 is more active as a transcrip-
fact that AL-438 generates a receptor conformation that
tional activator. Thus,AL-438 does not completely
differs from steroids,which in turn changes the spectrum of
separate transcriptional repression from activation,but
coactivators with which the receptor can interact. Certain
instead appears to be separating activities in a gene-speci®c
coactivators bind GR with identical af®nity in the presence
fashion. In animal models,AL-438 was as ef®cacious as
of either AL-438 or prednisolone,while others exhibit
prednisolone at inhibiting in¯ammation. Importantly,AL-
signi®cantly reduced af®nity in the presence of this com-
438 exhibited signi®cantly reduced impact on fasting
pound. We believe that since different genes have different
glucose levels compared with prednisolone,suggesting
requirements for speci®c coactivators,this may be the
that this compound might not cause the diabetogenic
molecular rationale for AL-438's gene-selective pro®le.
J.N. Miner / Biochemical Pharmacology 64 (2002) 355±361
and interleukin-6),but is weaker (a partial agonist) for
This is consistent with the gene-speci®c activity
transcriptional activation. Using other promoters in differ-
described earlier. The mechanism may be found in the
ent cell backgrounds,AL-438 is more active as a transcrip-
fact that AL-438 generates a receptor conformation that
tional activator. Thus,AL-438 does not completely
differs from steroids,which in turn changes the spectrum of
separate transcriptional repression from activation,but
coactivators with which the receptor can interact. Certain
instead appears to be separating activities in a gene-speci®c
coactivators bind GR with identical af®nity in the presence
fashion. In animal models,AL-438 was as ef®cacious as
of either AL-438 or prednisolone,while others exhibit
prednisolone at inhibiting in¯ammation. Importantly,AL-
signi®cantly reduced af®nity in the presence of this com-
438 exhibited signi®cantly reduced impact on fasting
pound. We believe that since different genes have different
glucose levels compared with prednisolone,suggesting
requirements for speci®c coactivators,this may be the
that this compound might not cause the diabetogenic
molecular rationale for AL-438's gene-selective pro®le.