L’ivermectine (Stromectol) est un antiparasitaire dont l’action repose sur la liaison sélective aux canaux chlore activés par le glutamate présents dans les cellules nerveuses et musculaires des parasites. Cette fixation entraîne une augmentation du flux de chlore, provoquant une hyperpolarisation et une paralysie irréversible. L’ivermectine est active contre la gale, l’onchocercose et certaines strongyloïdoses. Sa biodisponibilité orale est variable, augmentée par la prise alimentaire, et son élimination est principalement fécale via un métabolisme hépatique. Elle ne traverse pas la barrière hémato-encéphalique, ce qui limite les effets neurologiques chez l’homme. Les précautions concernent l’interaction avec les inhibiteurs du CYP3A4, ainsi que les réactions inflammatoires dues à la destruction massive des parasites. Dans les documents de référence, stromectol prix est associé à des protocoles précis adaptés aux différentes infestations, avec une attention particulière sur la sécurité d’emploi en cas d’immunodépression.
4w04t00s974p
Glutamate and Glutamine in Metabolism Glutamine and Glutamate Exchange between the Fetal Liver and the Placenta1 University of Colorado Health Sciences Center, Aurora, CO
The transport and metabolism of glutamine (GLN) and glutamate (GLU) during fetal development
exhibit unique characteristics that clearly emphasize the importance of the interaction between the placenta andthe fetal liver. GLN is delivered into the fetal circulation at a rate that is the highest of all the amino acids. In contrast,ϳ90% of fetal plasma GLU is extracted by the placenta. Conversely, the fetal liver has a large net output of GLUand a net uptake of GLN. We have studied the fluxes of GLU and GLN into and out of the placenta and fetal liver,as well as their interconversion in these organs, during late gestation in sheep. In the fetus, 45% of GLN carbontaken up by the liver exits as GLU; indeed, the production of GLU from GLN is large, ϳ3.7 mol/(min⅐kg fetus), andaccounts for virtually all of the GLU produced in the fetus. In contrast, only 6% of GLU carbon is converted to GLNin the placenta; most of the fetal plasma GLU taken up by this organ is converted to CO. Remarkably, placental
GLU uptake accounts for Ͼ60% of the fetal plasma GLU disposal rate. In some respects, the net output of GLUfrom the liver in fetuses replaces the net hepatic glucose output that is characteristic of postnatal life. We alsoexamined GLN and GLU fluxes in pregnant sheep during either dexamethasone-induced or spontaneous partu-rition. At parturition, a striking reduction in GLU output from the fetal liver occurred, leading to a fall in fetal arterialGLU concentrations and a marked decrease in placental GLU uptake. These changes were progressive asparturition advanced and correlated with a marked decrease in progesterone output from the pregnant uterus. J. Nutr. 130: 974S-977S, 2000.
KEY WORDS: ● placental uptake ● fetal liver ● glutamate ● glutamine ● parturition
Despite the evidence collected in adults that glutamine
quently confirmed in late-gestation ovine fetuses, both in our
(GLN)2 and glutamate (GLU) play unique roles in nutrition
laboratory and in those of others (Chung et al. 1998, Lemons
and metabolism, their functions during early development
and Schreiner 1984, Marconi et al. 1989). In addition, studies
have received scant attention. In fact, only about 20 years ago,
in rhesus monkeys of the transport of labeled GLU from the
while studying the umbilical uptake of nutrients by the ovine
maternal into the fetal circulation demonstrated there to be
fetus, did we make the initial, key observation that the pla-
little or no GLU transport across the primate placenta (Ste-
centa takes up GLU from the fetal circulation, while concur-
gink et al. 1975). Others have shown in humans during ce-
rently releasing GLN into the fetal circulation in very large
sarean section (when both umbilical arterial and venous blood
amounts (Lemons et al. 1976). From this finding, it was clear
samples can be obtained) that the fetus demonstrates a nega-
that all fetal GLU requirements must be met by the fetal
tive (umbilical vein Ϫ fetal artery) concentration difference
for GLU across the placenta (Hayashi et al. 1978). Thisfinding confirms in humans, as in other species, that there is a
Net fluxes of glutamine and glutamate
net uptake of GLU from the umbilical circulation into the
The early observation that GLU is extracted from the fetal
placenta. Hence, this phenomenon is not unique to the epi-
circulation by the placenta (Lemons et al. 1976) was subse-
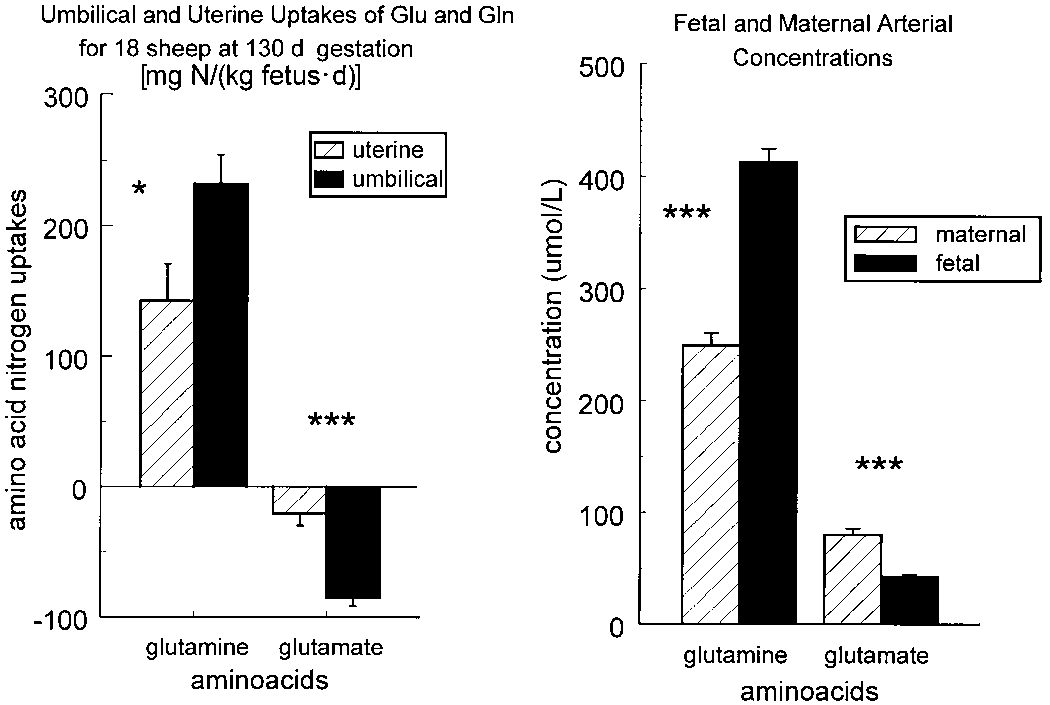
theliochorial placenta, but seems to be a more general char- acteristic of trophoblasts. Figure 1 presents data from a recent study of 18 pregnant sheep, summarizing the umbilical and
1 Presented at the International Symposium on Glutamate, October 12-14,
uterine uptakes of GLN and GLU (Chung et al. 1998). Note
1998 at the Clinical Center for Rare Diseases Aldo e Cele Dacco', Mario Negri
that GLU is taken up by the placenta from both circulations.
Institute for Pharmacological Research, Bergamo, Italy. The symposium was
Additionally, GLN delivery to the fetus (i.e., its umbilical
sponsored jointly by the Baylor College of Medicine, the Center for Nutrition at theUniversity of Pittsburgh School of Medicine, the Monell Chemical Senses Center,
uptake) is significantly greater than uterine uptake, demon-
the International Union of Food Science and Technology, and the Center for
strating net placental GLN production. In the 1980s, fetal
Human Nutrition; financial support was provided by the International Glutamate
surgery progressed to a point that permitted sampling of the
Technical Committee. The proceedings of the symposium are published as asupplement to The Journal of Nutrition. Editors for the symposium publication
venous drainage from the fetal liver. The preparation we
were John D. Fernstrom, the University of Pittsburgh School of Medicine, and
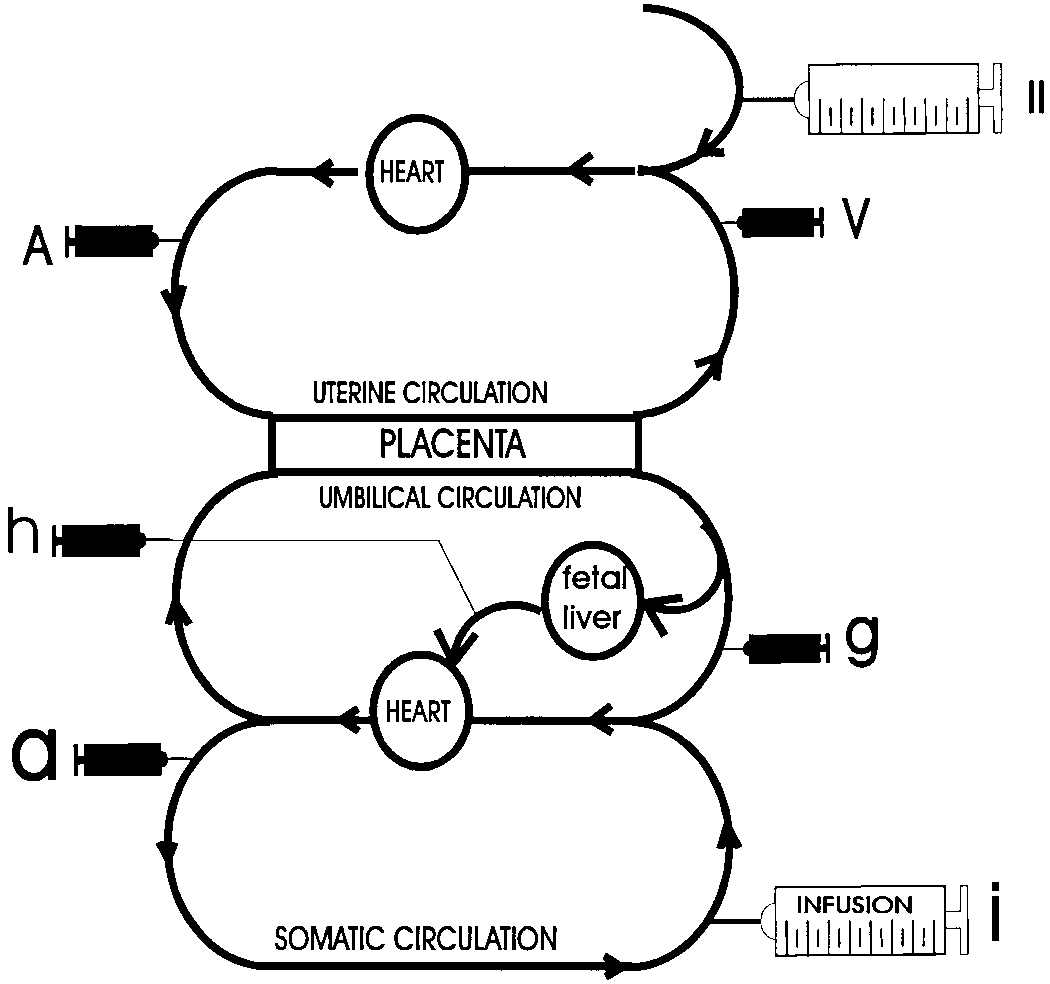
utilized is described in Figure 2, with potential infusion sites
Silvio Garattini, the Mario Negri Institute for Pharmacological Research.
for tracers in both the maternal and fetal circulations. Thus,
Abbreviations used: BCAA, branched-chain amino acid; GLN, glutamine;
for the first time, we were able to look at the fluxes of amino
0022-3166/00 $3.00 2000 American Society for Nutritional Sciences.
The uterine and umbilical uptakes of glutamate (Glu)
and glutamine (Gln) are presented as well as their fetal and maternal
The net fluxes, measured in vivo, of the branched-
arterial concentrations. The uptakes for each circulation were calcu-
chain amino acids, glutamine, glutamate and ammonia, into and out of
lated as the (flow ϫ arteriovenous concentration difference). *P Ͻ 0.05,
the ovine placenta. The values are expressed in mol/kg fetus/min.
***P Ͻ 0.001 (paired t test). From Chung et al. (1998).
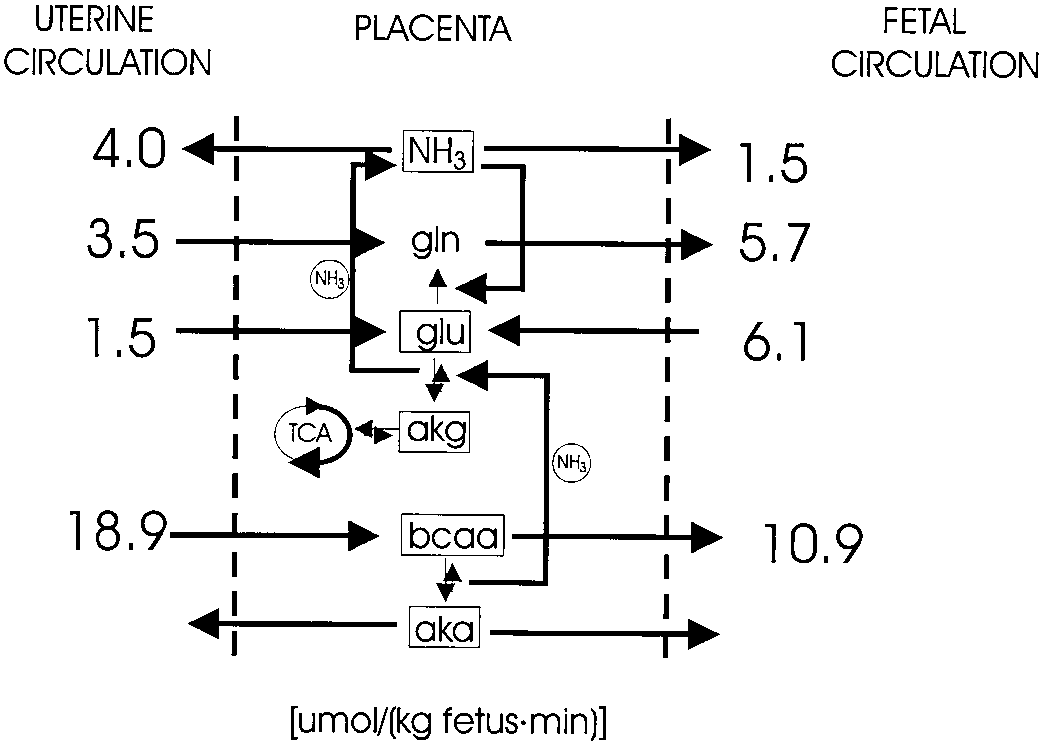
Note the contribution of the branched-chain amino acids to both glu-tamate and NH production within the placenta. Abbreviations: gln,
glutamine; glu, glutamate; akg, ␣-ketoglutarate; TCA, tricarboxylic acidcycle; bcaa, branched-chain amino acids; aka, branched-chain ␣-keto
acids into and out of the fetal liver and placenta simulta-
acids; NH , ammonia. From Chung et al. (1998), Loy et al. (1990), and
neously. Subsequent studies using this procedure revealed the
existence of important interorgan cycles for amino acids be-tween fetal liver and placenta. Specifically, we observed theopposite arrangement for GLU and GLN across the fetal liverthan that across the placenta. That is, the fetal liver experi-
Placental glutamate supply
ences a large uptake of GLN from the fetal circulation, and a
Because there is little uterine uptake of GLU, placental
large net hepatic release of GLU, a phenomenon that is not
GLU supply is determined by measuring placental GLU pro-
found in normal postnatal hepatic metabolism. In essence, we
duction and GLU delivery to the placenta from the fetal
found the following: 1) the placenta delivers GLN into the
circulation. The coefficient of extraction of GLU from fetal
fetal circulation; 2) GLN is extracted by the fetal liver and
plasma as it perfuses the placenta is ϳ90%, a very high value
used for the net hepatic release of GLU; and 3) the GLU
that is unique to GLU (Moores et al. 1994). Thus, the GLU
circulating in fetal blood is taken up by the placenta.
supply to the placenta is determined primarily by the umbilicaldelivery rate (represented by the umbilical plasma flow) ϫ the
fetal arterial GLU concentration. The latter is a function offetal hepatic GLU release. Tracer GLU and GLN studies ofthe fetal lamb have shown that the hepatic production rate ofglutamate from glutamine is virtually identical to the total fetalglutamate production rate from glutamine (Vaughn et al. 1995). Thus, the fetal liver is the primary site for glutamateproduction and, as such, also determines the glutamate supplyto the placenta.
Recent data from our laboratory suggest that the placental
production of GLU from oxoglutarate may be driven by thehigh rate of transamination of the branched-chain amino acids(BCAA) to their respective keto acids. The ovine placentahas a high level of activity of the branched-chain transami-nases, which is consistent with other data on tracer leucinefluxes across the placenta and in the fetal circulation. Thesestudies have shown that ϳ20 -25% of leucine uptake from the
maternal circulation is utilized within the placenta (Loy et al. 1990). The nitrogen derived from the metabolism of BCAAinto their respective keto acids contributes to both placentalNH
production and GLU formation from oxoglutarate
(Jo'zwik et al. 1999). Thus, the placental supply of GLU derives from both its uptake from the fetal circulation and its production in the placenta associated with BCAA transami- nation. Figure 3 summarizes data from several studies (Chung
Schematic of the infusion and sampling sites utilized
et al. 1998, Jo'zwik et al. 1999, Loy et al. 1991) and indicates
for tracer studies in late gestational lambs (see text). Abbreviations: A,
the net uptake or release from sheep placenta of the BCAA,
maternal artery sample; V, uterine vein sample; a, umbilical arterysample; g, umbilical vein sample; h, left fetal hepatic vein; i, fetal venous
GLN and GLU into the uterine and umbilical circulations.
infusion; II, maternal venous infustion.
When L-[1-14C] GLU is infused into the fetal circulation,
uptake from the fetal plasma (arteriovenous differences acrossthe umbilical circulation fell from control values of 18 Ϯ 3 to2 Ϯ 3 mol/mmol O ). At the same time, progesterone output
from the pregnant uterus also decreased significantly. Thus,the events leading up to parturition are associated with pro-found changes in fetal hepatic and placental GLU and GLNmetabolism. However, with the use of this paradigm, we couldnot distinguish whether these changes were due to the manyendocrine changes associated with parturition or simply to thedexamethasone used to induce parturition.
Our ongoing studies are attempting to clarify this latter
issue, but at present are very preliminary. One study examinedfetal hepatic and placental GLU and GLN metabolism duringspontaneous parturition (Timmerman et al., unpublished ob-servations). The experimental design enables us to sample thefetal circulation, including the hepatic venous circulation and
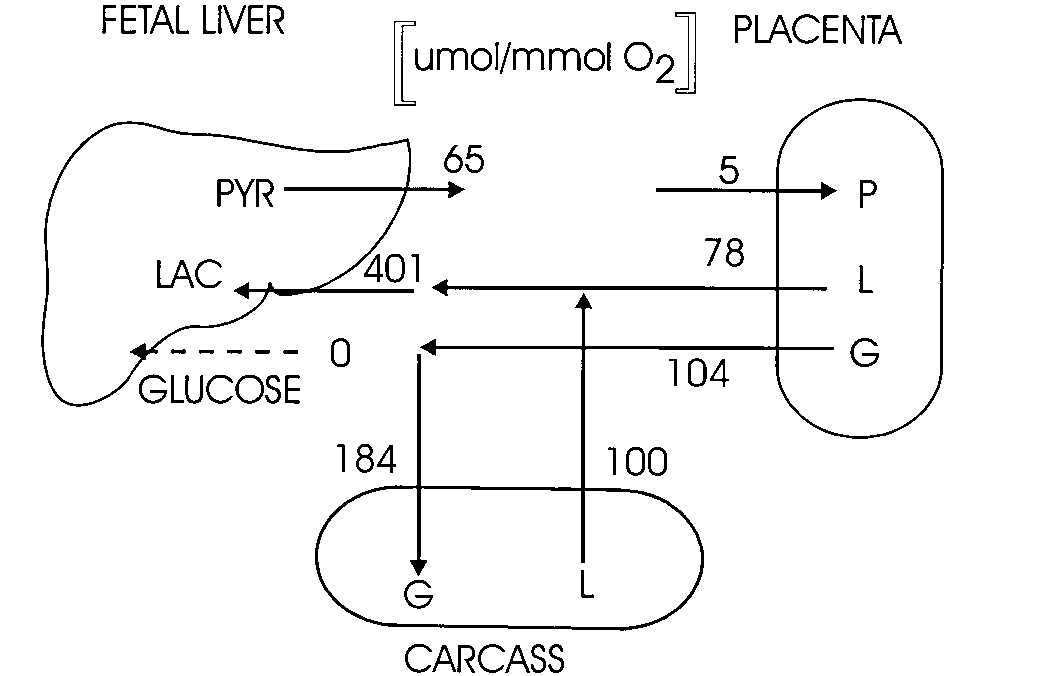
The exchange of glucose, lactate and pyruvate among
the liver, placenta, and carcass in the fetal lamb. The data for the
the maternal uterine circulation, beginning 7-10 d before
carcass are calculated from measured values for the fetal hindlimb.
expected parturition. The results to date have revealed both
PYR ϭ P ϭ pyruvate, LAC ϭ L ϭ lactose, G ϭ glucose. [Timmerman et
similarities to and differences from dexamethasone-induced
al., unpublished observations, and Wilkening et al. (1994)].
parturition. The similarities relate to changes in GLU andprogesterone metabolism. During spontaneous parturition,there is a marked decrease in net fetal hepatic GLU output,
ϳ80% of the carbon can be accounted for as CO , half in the
leading to a decrease in placental GLU uptake from the fetal
fetus and half in the placenta (Moores et al. 1994). When
circulation (see Fig. 5, which presents data for a single ani-
tracer-labeled GLN is infused into the fetal circulation, ϳ50%
mal). Coincident with these changes, progesterone output
of the hepatic uptake exits the fetal liver as GLU (Vaughn et
al. 1995). The calculated fetal hepatic production rate of GLU
A second preliminary study examined whether GLN car-
from GLN is 3- 4 mol/(kg fetus⅐min). Both GLN and GLU
bon flux within the fetal liver is altered during parturition
(and alanine) are taken up by fetal hindlimb tissues (Wilken-
(Timmerman et al., unpublished observations). We utilized
ing et al. 1994). The fact that the fetal carcass, as represented
the model of dexamethasone-induced parturition to study the
by the hind limb tissues, takes up both GLN and alanine is
fluxes of L-[1-13C] GLN and L-[3H ϩ3H ] GLU in the fetal
consistent with data showing there to be no significant rate of
circulation. These fluxes were measured in each animal before
fetal hepatic gluconeogenesis (Hay et al. 1984). GLU delivery
and after a 25-h fetal infusion of dexamethasone. The most
to the carcass and placenta comes primarily from the fetal liver
significant finding was that the ratio of 13CO to GLU
and indirectly via GLN delivery from the placenta.
leaving the fetal liver was significantly higher during the
In one sense, the large GLU output from the fetal liver can
dexamethasone infusion compared with a control period.
be equated with the large hepatic glucose output during post-
Thus, GLN carbon is redirected into oxoglutarate and the
natal life. We have shown that there is no significant gluconeo-
tricarboxylic acid cycle and away from GLU release.
genesis nor any significant glucose output from the fetal liver during normal gestation (Hay et al. 1984). This is presumably useful to the fetus because fetal glucose production would block the transplacental transport of glucose from the mater- nal to the fetal circulation. Figure 4 summarizes the carbohy- drate exchange among the fetal liver, placenta and carcass taken from recent data (Timmerman et al., unpublished re- sults) and Wilkening et al. (1994). Changes in glutamine-glutamate metabolism during parturition
During parturition, endocrine changes occur in the fetal
circulation that signal a shift from the fetal to the postnatalpattern of net hepatic glucose or GLU release. For this reason,we thought it would be instructive to study net hepatic andplacental uptake and/or release of GLN and GLU around thetime of parturition. To facilitate these studies, we used a fetalinfusion of dexamethasone to induce labor in late-gestationalfetal lambs (Barbera et al. 1997). The arteriovenous concen-tration differences for GLN and GLU were measured in acontrol period that preceded dexamethasone infusion, andthen at 25 h and at 40 - 48 h after dexamethasone infusionbegan. At 25 h, GLU release from the fetal liver had fallen
Changes in fetal hepatic and placental glutamate (glu)
dramatically from 180 Ϯ 56 to 45 Ϯ 18 mol/mmol O. This
uptake before parturition. The data are derived from a single animal for
change produced a significant fall in fetal plasma GLU con-
GLU release from the fetal liver and GLU uptake by the placenta.
centrations and led to a significant decline in placental GLU
(Timmerman et al., unpublished observations).
amino acids and aminonitrogen following maternal infusion of amino acidsinto pregnant sheep. Am. J. Obstet. Gynecol. 180: 447- 453.
Lemons, J. A. & Schreiner, R. L.
Glutamine and GLU metabolism play important and
during the fed and fasted states. Ann. Nutr. Metab. 28: 268 -280.
unique roles during fetal development. Their interorgan ex-
Lemons, J. A., Adcock, E. W., III, Jones, M. D., Jr., Naughton, M. A., Meschia, G.
change (between fetal liver and placenta) and particularly, the
Umbilical uptake of amino acids in the unstressed
fetal lamb. J. Clin. Investig. 58: 1428 -1434.
fetal liver's central role in maintaining GLU supply to the
Loy, G. L., Quick, A. N., Jr., Battaglia, F. C., Meschia, G. & Fennessey, P. V.
placenta, illustrate that these two organs form an integrated
Measurement of leucine and ␣-ketoisocaproic acid fluxes in the
fetal/placental unit. J. Chromatogr. Biomed. Appl. 562: 169 -174.
Loy, G. L., Quick, A. N., Jr., Hay, W. W., Jr., Meschia, G., Battaglia, F. C. &
Feto-placental deamination and decarboxylation
of leucine. Am. J. Physiol. 259: E492-E497. LITERATURE CITED
Marconi, A. M., Battaglia, F. C., Meschia, G. & Sparks, J. W.
parison of amino acid arteriovenous differences across the liver, hindlimb and
Barbera, A., Wilkening, R., Battaglia, F. C. & Meschia, G.
placenta in the fetal lamb. Am. J. Physiol. 257: E909 -E915.
alterations in the fetal hepatic and umbilical circulations during glucocorticoid
Moores, R. R., Jr., Vaughn, P. R., Battaglia, F. C., Fennessey, P. V., Wilkening,
induced parturition in sheep. Pediatr. Res. 41: 242-248.
Glutamate metabolism in the fetus and placenta
Chung, M., Teng, C., Timmerman, M., Meschia, G. & Battaglia, F. C.
of late gestation sheep. Am. J. Physiol. 267: R89 -R96.
Production and utilization of amino acids by ovine placenta in vivo. Am. J.
Stegink, L. D., Pitkin, R. M., Reynolds, W. A., Filer, L. J., Jr., Boaz, D. P. &
Placental transfer of glutamate and its metabolites
Hay, W. W., Jr., Sparks, J. W., Wilkening, R. B., Battaglia, F. C. & Meschia, G.
in the primate. Am. J. Obstet. Gynecol. 122: 70 -78.
Fetal glucose uptake and utilization as functions of maternal glucose
Vaughn, P. R., Lobo, M., Battaglia, F. C., Fennessey, P., Wilkening, R. & Meschia,
concentration. Am. J. Physiol. 246: E237-E242.
Glutamine-glutamate exchange between placenta and fetal liver.
Hayashi, S., Sanada, K., Sagama, N., Yamada, N. & Kido, K.
vein-artery differences of plasma amino acids in the last trimester of human
Wilkening, R. B., Boyle, D. W., Teng, C., Meschia, G. & Battaglia, F. C.
pregnancy. Biol. Neonate 34: 11-18.
Amino acid uptake by the fetal ovine hind limb under normal and euglycemic
Jo'zwik, M., Teng, C., Battaglia, F. C, & Meschia, G.
hyperinsulinemic states. Am. J. Physiol. 266: E72-E78.
Medicatieprotocol behorend bij het onderzoek P-CGT +/- AD bij recidiverende depressie Inleiding In dit onderzoek wordt het effect van P-CGT plus doorgaan met antidepressiva (AD) vergeleken met enerzijds doorgaan met AD plus care as usual en anderzijds P-CGT plus afbouwen van AD Een eerste probleem is dat de farmacotherapiecondities in alle armen zoveel mogelijk uniform moet geb
Working Guidelines Sarah MATHESON and John OSHA, Deputy Reporters General Anne Marie VERSCHUR, Sara ULFSDOTTER and Kazuhiko YOSHIDA Second medical use and other second indication claims Introduction This question seeks to determine the type, scope and enforcement of patent protection for new uses of known chemical compounds when a known substance is found to have a new therapeutic
 The uterine and umbilical uptakes of glutamate (Glu)
and glutamine (Gln) are presented as well as their fetal and maternal
The net fluxes, measured in vivo, of the branched-
arterial concentrations. The uptakes for each circulation were calcu-
chain amino acids, glutamine, glutamate and ammonia, into and out of
lated as the (flow ϫ arteriovenous concentration difference). *P Ͻ 0.05,
the ovine placenta. The values are expressed in mol/kg fetus/min.
The uterine and umbilical uptakes of glutamate (Glu)
and glutamine (Gln) are presented as well as their fetal and maternal
The net fluxes, measured in vivo, of the branched-
arterial concentrations. The uptakes for each circulation were calcu-
chain amino acids, glutamine, glutamate and ammonia, into and out of
lated as the (flow ϫ arteriovenous concentration difference). *P Ͻ 0.05,
the ovine placenta. The values are expressed in mol/kg fetus/min.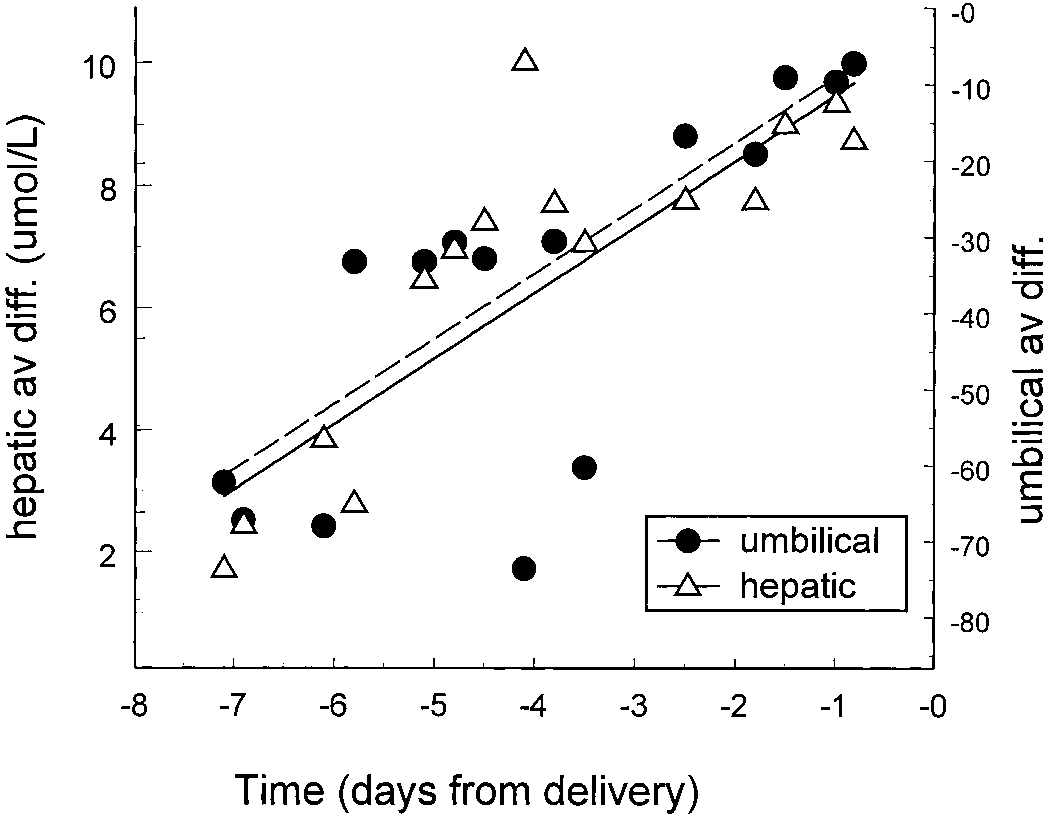
 uptake from the fetal plasma (arteriovenous differences acrossthe umbilical circulation fell from control values of 18 Ϯ 3 to2 Ϯ 3 mol/mmol O ). At the same time, progesterone output
from the pregnant uterus also decreased significantly. Thus,the events leading up to parturition are associated with pro-found changes in fetal hepatic and placental GLU and GLNmetabolism. However, with the use of this paradigm, we couldnot distinguish whether these changes were due to the manyendocrine changes associated with parturition or simply to thedexamethasone used to induce parturition.
uptake from the fetal plasma (arteriovenous differences acrossthe umbilical circulation fell from control values of 18 Ϯ 3 to2 Ϯ 3 mol/mmol O ). At the same time, progesterone output
from the pregnant uterus also decreased significantly. Thus,the events leading up to parturition are associated with pro-found changes in fetal hepatic and placental GLU and GLNmetabolism. However, with the use of this paradigm, we couldnot distinguish whether these changes were due to the manyendocrine changes associated with parturition or simply to thedexamethasone used to induce parturition.